


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Рост нитчатых талломов может происходить по-разному. Различают диффузный рост (делятся все клетки), апикальный (делятся преимущественно конечные клетки), интеркалярный (зона роста расположена в средней части нити, обычно ближе к вершине) и базальный рост (делятся клетки в основании нити).
Возникновение нитчатого типа структуры было качественно новым этапом в развитии растительного мира. Он послужил отправным пунктом для развития других, более сложных типов структуры.
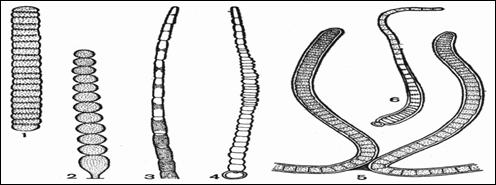

1 – нить с диффузным ростом у Oscillatoria;
2 – нить с дифференцированным основанием у Endonema;
3, 4 – верхушка нити у Rivularia, вытянутая в волосок;
5 – апикальный рост на концах ветвей у Scytonema;
6 – базальный рост у Calothrix.
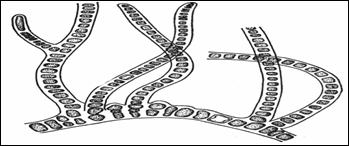
Разнонитчатая (или Гетеротрихальная) организация представляет собой нитчатую структуру с морфологически и функционально дифференцированными многоклеточными участками. Чаще всего разнонитчатый таллом состоит из двух частей – стелющейся по субстрату и прямостоящей (слайд). Горизонтальная часть таллома состоит или из различимых нитей, или они полностью смыкаются в сплошную пластинку. В этом случае она нарастает по периферии и уже не имеет следов нитчатого строения. Развитие вертикальной и горизонтальной частей бывает разным, вплоть до редукции какой-либо части. Если редуцируется горизонтальная часть, то таллом становится сходным с нитчатым. Если исчезают вертикальные части, то остается лишь простой однослойный диск, целиком прикрепленный к субстрату, имеющий сходство со следующим, пластинчатым типом строения, но отличающимся по способу формирования.
У некоторых водорослей вертикальные нити дифференцированы на междоузлия и узлы, от которых отходят мутовки боковых ветвей. Этот тип структуры таллома распространен у зеленых, бурых и красных водорослей.

Пластинчатая (или тканевая) структура характеризуется многоклеточными слоевищами в форме пластинок (слайд). Состоят они из одного или нескольких слоев клеток. Такие талломы могут иметь сложную форму.
Образуются такие пластинки из нити в результате продольного деления клеток. Если клетки делятся только в одной плоскости, получается однослойное слоевище, если в двух – 2-многослойное. Разновидность пластинчатой структуры – трубчатое слоевище. Образуется оно из двухслойных талломов в результате расхождения слоев в середине при сохранении связи по краям. При этом внутри образуется полость, окруженная однослойными стенками.
Пластинчатый тип слоевища может ветвиться. Он растет либо распростертый по субстрату, или прикрепленный краем пластинки. Пластинчатые слоевища хорошо представлены в отделах зеленых, бурых и красных водорослей.
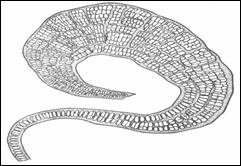

Пластинчатая структура у зеленой водоросли Prasiola:
Однорядная нить, разрастающаяся в однослойную пластинку
Харофитная структура свойственна только харовым водорослям. Их структура характеризуется наличием главной оси, мутовками членистых боковых осей и отходящими к низу ризоидами. Места расположения мутовок называют узлами, части главной оси между ними – междоузлиями. Междоузлия и членики боковых осей образованы одиночной вытянутой клеткой, но у ряда видов рода хара эти клетки снаружи покрыты слоем других клеток, образующих кору. Такой принцип членисто-мутовчатого строения растительного организма повторяется на более высоком уровне организации у хвощей и некоторых цветковых растений (род казуарина).
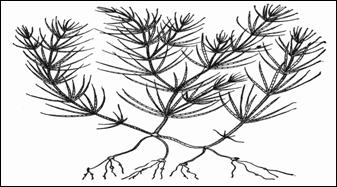

Молодой экземпляр Chara
Сифонокладальная форма представлена талломами с многоядерными клетками.
Сифональную (или сифоновую) структуру часто называют неклеточной. Организмы отличаются отсутствием клеточных перегородок и талломы как бы представляют собой одну клетку часто с большим количеством ядер. При этом размеры организма бывают довольно большими и сильно расчлененными внешне. Сифональная структура хорошо выражена в отделах зеленых и желто-зеленых водорослей.

Сифональная структура у морской зеленой водоросли Caulerpa (часть таллома)
Перечисленные ступени организации талломов в разных отделах представлены неодинаково. Наиболее полным спектром форм характеризуются зеленые, золотистые, желтозеленые и динофитовые водоросли. Крайне интересно, что цианобактерии, которые являются прокариотами, также демонстрируют разные типы морфологической структуры таллома. Следует отметить, что у многоклеточных форм, гаметы и споры имеют монадный и коккоидный тип структуры. У высокоорганизованных форм разные стадии развития могут иметь разный тип структуры таллома.
Если расположить некоторые роды водорослей в таблицу, в которой вертикальные ряды отличаются по окраске талломов, а горизонтальные – по их морфологии, то получим сеть наподобие периодической системы элементов . Этот морфологический параллелизм отражает близкие пути эволюции отдельных групп водорослей. Это фактически классификация водорослей, сложившаяся в 19 - первой половине 20 века. В качестве основных признаков использовали как морфологию (выделяли классы и отделы по горизонтальным рядам), так и пигментацию, проводя разделение по вертикальным столбцам.


Главная причина этого параллелизма – сходство образа жизни и способов питания. Сходный образ жизни обуславливает сходные каналы эволюции. Значительную роль в формировании разнообразия живых организмов сыграл также первичный и вторичный эндосимбиоз, который привел к возникновению качественно иных организмов.
Симбиоз – это различные формы совместного существования разных организмов. Здесь я использую термин симбиоз в узком смысле, соответствующем понятию «мутуализм». Мутуализм – это форма симбиоза, при которой два различных организма возлагают друг на друга регуляцию своих отношений с внешней средой. При этом отношения между партнерами взаимно выгодны и один из них не может существовать без другого. «Далеко зашедший» симбиоз можно рассматривать как особый способ эволюции, при котором сложные организмы образуются из нескольких простых. Теория о происхождении организмов путем симбиоза называется симбиогенез.
Теория эндосимбиоза описывает происхождение клеточных органелл от различных групп прокариот, в частности, пластид от подобных цианобактериям прокариот, живущих в эукариотных клетках. Эндосимбионты в значительной степени редуцированы, но морфологические, биохимические данные, данные молекулярной биологии во многом указывают на прокариотов как на предков пластид. Первичный эндосимбиоз дал начало пластидам с двумя ограничивающими мембранами. Это хлоропласты зеленых водорослей, высших растений, красных водорослей.
В следующем витке эндосимбиоза принимали участие только эукариоты. Красные или зеленые водоросли были проглочены и сохранены другими эукариотами. Таким образом был приобретена возможность фотосинтеза у других эволюционных групп. Это вторичный эндосимбиоз, в результате которого получились пластиды, имеющие три или четыре ограничивающие мембраны. В результате у зеленых и красных водорослей двухслойная оболочка хлоропласта, у динофитовых и эвгленовых – 3-х слойная, у золотистых, желтозеленых, диатомовых, криптофитовых и бурых – четырехмембранная. При вторичном эндосимбиозе захваченная клетка значительно редуцировалась до хроматофора и окружающих мембран, которые возникли как результат захвата. Для динофитовых есть основания предполагать третичный эндосимбиоз. Получается, что клетки некоторых организмов, подобно матрешкам, содержат вставленные друг в друга мелкие клетки.
Вегетативное, бесполое и половое размножение водорослей и их формы вы изучите самостоятельно. Эти вопросы хорошо рассмотрены в «Жизни растений» и «Курсе низших растений».
ЦИКЛЫ РАЗВИТИЯ ВОДОРОСЛЕЙ.
Изменения, претерпеваемые особями вида между одноименными стадиями, составляют его цикл развития. Жизненные циклы в наиболее полном виде включают следующие фазы развития: вегетативный рост, бесполые спороношения, половой процесс, покоящиеся стадии.
В зависимости от места мейоза в жизненном цикле различают следующие его виды:
Типы мейоза:
1. гаметический – гаплоидные гаметы формируются на диплоидном талломе (Fucus и некоторые другие бурые водоросли, диатомовые); характерна для животных. Вся вегетативная фаза диплоидна
2.спорический – гаплоидные споры бесполого размножения формируются на диплоидном талломе (бурая водоросль Laminaria);
3. зиготический – при прорастании зиготы (зеленые водоросли: вольвоксовые, хлорококковые);
4. соматический – гаплоидные вегетативные клетки формируются на диплоидном талломе; впоследствии из них образуются гаметы (зеленая водоросль Prasiola).
Типы жизненных циклов водорослей:
1. Бесполый. Половой процесс отсутствует. Размножение осуществляется гаплоидными спорами Таллом гаплоидный, диплоидной стадии нет (зеленые водоросли: хлорококковые, эвгленовые, криптофитовые и др.)
2. Гаплоидный. На гаплоидном талломе формируются споры и гаметы. После слияния гамет образуются покоящиеся зиготы. Вегетативное тело гаплоидно, диплоидна лишь зигота. Мейоз зиготический. (многие зеленые водоросли).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 |
