


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Наиболее распространены из диатомей:
- Навикула (Navicula), створки лодочкообразные с острыми или суженными концами.
- Пиннулярия (Pinnularia), створки удлиненно-элиптические со швом и хорошо заметной штриховатостью.
- Цимбелла (Cymbella), створки серповидно-изогнутые.
Навикула, пиннулярия и цимбелла являются бентосными водорослями, относятся к классу Пеннатных. Из центрических планктонных можно встретить в наших водоемах циклотеллу (Cyclotella), одиночные клетки которой выглядят в виде низкой круглой коробочки.
Отдел бурые водоросли – Phaeophyta.
К отделу бурых водорослей (около 1500 видов) относятся многочисленные макроскопические водоросли, общим внешним признаком которых служит желтовато-бурая окраска их слоевищ, обусловленная наличием желтых и бурых пигментов. Преимущественно дисковидные хлоропласты содержат хлорофиллы а и с, каротины и ксантофиллы. Пиреноиды очень мелкие. Запасные питательные вещества - ламинарин, маннит (сахароспирт), в небольших количествах - жир. Кроме обычных органелл в клетках присутствуют физоды, структуры, содержащие танины. Оболочки клеток снаружи ослизняющиеся - пектиновые, внутренний слой - целлюлоза.
Тип структуры таллома бурых водорослей преимущественно пластинчатый, нередко очень сложно организованный, редко разнонитчатый. Подавляющее большинство представителей этой группы – морские формы.
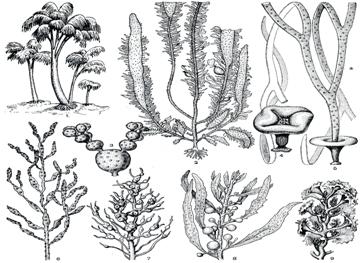



Слоевища бурых водорослей бывают размером от нескольких десятков микрометров до десятков метров (макроцистис – до 50 м). У высокоорганизованных форм талломы дифференцированы и напоминают цветковые растения. Некоторые крупные представители имеют воздушные пузыри. Все бурые водоросли растут прикрепленными к грунту или другим водорослям. Для прикрепления они формируют ризоиды или базальный диск. На поперечном разрезе крупных талломов бурых водорослей различаются зоны, занятые клетками разного строения, специализированными для выполнения отдельных функций.
Клетки одноядерные, одетые толстыми оболочками с крупными порами. Оболочка состоит из внутреннего целлюлозного и внешнего слоя, основу которого составляют белки, соединенные с альгиновой кислотой и ее солями. Альгулоза. Физоды. Хлоропласты обычно мелкие дисковидные, реже – лентовидные и пластинчатые. У ряда таксонов в хлоропластах имеется пиреноид. В хлоропластах тиллакоиды группируются в ламеллы по 3. Пигменты: хлорофилл a, c, каротиноиды. Запасные питательные вещества в клетках вне хлоропластов - ламинарин (полисахарид) и маннит (шестиатомный спирт), в небольших количествах липиды.
Размножение. У Б. В. встречаются все основные типы размножения – вегетативное, бесполое и половое. Вегетативное размножение происходит при случайном отделении ветвей от слоевища, у этих ветвей не образуются органы прикрепления и не формируются органы бесполого и полового размножения. Органы размножения образуются на них в случае, если их развитие к моменту отрыва слоевища от грунта уже началось. Бесполое размножение осуществляется зооспорами зооспорами (есть виды, размножающиеся моно - или тетраспорами). Строение зооспор. Зооспоры грушевидной формы имеют сбоку два неравных жгутика: передний - длинный, задний - короткий. Образование зооспор и гамет у бурых водорослей происходит во вместилищах двух основных типов: одногнездных и многогнездных. Мейоз происходит при формировании спор в одногнездных спорангиях. Половой процесс изогамный, гетерогамный и оогамный. Все бурые водоросли, за исключением фукусовых, имеют чередование поколений; на бесполом поколении (спорофите) в зооспорангиях (или тетраспорангиях) после редукционного деления образуются зооспоры (или тетраспоры); из них вырастают гаплоидные половые растения (гаметофиты) раздельнополые или обоеполые. После оплодотворения зигота без периода покоя развивается в новое бесполое диплоидное растение (спорофит). У фукусовых вся жизнь водоросли происходит в диплоидной фазе, гаплоидны лишь сперматозоиды и яйцеклетки, перед образованием которых происходит редукционное деление.
В зависимости от характера чередования поколений и смены ядерных фаз бурые водоросли делят на 3 класса.
Класс Изогенератные (Isogeneratae)
У большинства водорослей этого класса спорофит и гаметофит одинаковы по форме и величине или не очень резко отличаются по величине. Половой процесс изо-, гетеро - или оогамный. Широко распространены в морях представители рода эктокарпус (Ectocarpus); особенно много их в холодных морях. Произрастают в литорали и сублиторали. Участвуют в обрастании судов, буйков. Они имеют вид маленьких кустиков или дерновинок, состоящих из обильно ветвящихся однорядных нитей, оканчивающихся нередко бесцветными многоклеточными волосками. Рост нитей интеркалярный. Сбоку на ветвях образуются одноклеточные зооспорангии. В них происходит редукционное деление и еще ряд делений клеточного ядра и формируется
много зооспор, которые выходят из зооспорангия, недолго поплавав, прорастают в такое же по внешности, но гаплоидное растение. На нем, на коротких боковых веточках образуются многоклеточные гаметангии, в которых развиваются одинаковые по форме и величине, но отличающиеся по поведению (+ и -) гаметы. Зигота без периода покоя прорастает в новую диплоидную нить с зооспорангием. Для эктокарпуса характерна изоморфная смена поколений. Половой процесс - изогамия.
Кутлярия (Cutleria) - распространена у европейских берегов Атлантического океана и в Средиземном море. Она имеет вид дихотонически разветвленных многослойных лент до 20 см длины, оканчивающихся волосками. Внешне кутлериевые отличаются от других водорослей по наличию бахромы из нежных волосков на вершине или по краю слоевища. У основания волосков на одних экземплярах развиваются крупноклеточные макрогаметангии, дающие двужгутиковые макрогаметы, на других экземплярах - развиваются мелкоклеточные микрогаметангии, дающие микрогаметы. После оплодотворения зигота без периода покоя развивается в новое диплоидное растение - спорофит, имеющее форму многослойной пластинки или корочки, плотно прижатой к субстрату. Она настолько не похожа на гаметофит, что была описана под новым родовым названием аглаозонии (Aglaozonia). На верхней стороне ее развиваются одноклеточные зооспорангии, в которых после редукционного деления формируются зооспоры. Зооспоры прорастают в лентовидный кустистый гаметофит. Род кутлярия пользуется наибольшей известностью. Она служит примером гетероморфной смены поколения, половой процесс ее - гетерогамия.
Диктиота (Dictyota) произрастает преимущественно в тропических и субтропических морях, встречается и в Черном море. Для нее характерно вильчато-разветвленное слоевище с расположенными в одной плоскости ветвями. Высота растения около 20 см, ширина ветвей 4-8 мм. Диктиота имеет изоморфное чередование поколений. На спорофите из поверхностных клеток развиваются крупные шаровидные тетраспорангии, в которых после редукционного деления образуется 4 гаплоидные неподвижные голые тетраспоры. Из тетраспор развиваются мужские (с многочисленными антеридиями, дающими до 30000 сперматозоидов) и женские (с оогониями, продуцирующими по одной яйцеклетке) гаметофиты. Зрелые яйцеклетки выпадают из оогониев. После оплодотворения зигота развивается в новый спорофит. Диктиота - пример развития водоросли с изоморфной сменой поколений, у которой половой процесс - оогамия.
Класс Гетерогенератные (Heterogeneratae)
В цикле развития водорослей этого класса чередуются макроскопический (у многих мощно развитый) спорофит и микроскопический мелкий гаметофит, который нередко по аналогии с папоротниками называется заростком. Половой процесс изогамный или оогамный. Спорофиты разных родов резко различаются и представляют собой самые крупные низшие растения достигающие в длину 60-100 м со значительным морфологическим расчленением и сложным анатомическим строением. Спорофиты расчленяются на листовую пластинку, черешок и ризоиды или базальный диск. В месте перехода пластинки в черешок - интеркалярная меристема. Спорофит многолетний, за исключением пластинки, которая ежегодно отмирает и заменяется новой. В меристодерме пластинки перед ее разрушением образуются одноклеточные зооспорангии. В каждом зооспорангии после предварительного редукционного деления образуется от 16 до 64 (иногда 128) зооспор. На одном экземпляре водоросли формируются миллиарды зооспор. После недолгого периода движения зооспоры прорастают в микроскопически мелкие нитевидные мужские и женские гаметофиты (заростки). В антеридиях образуется по одному сперматозоиду, в оогониях - по одной яйцеклетке. После оплодотворения из зиготы развивается постепенно новый мощный спорофит.
Итак, для гетерогенаратных водорослей характерна гетероморфная смена поколений. Половой процесс - оогамия.
Ламинария (Laminaria), виды рода широко распространены в северных морях. Таллом расчленен на листовую пластинку, ствол и ризоиды. Встречается по вертикали от поверхности моря до глубины 200 м.
Продольный разрез черешка молодого слоевища ламинарии
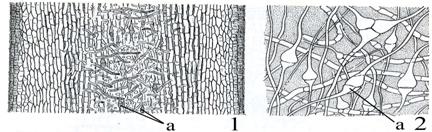
 1 – при малом увеличении, 2 – при большом увеличении: а – трубчатые нити.
1 – при малом увеличении, 2 – при большом увеличении: а – трубчатые нити.
Макроцистис (Macrocystis), растущий на глубине 10-20 м, имеет ствол многократно разветвленный до 50-60 м длиной. Перед каждой пластинкой ветвь обычно расширена в грушевидный воздушный пузырь. Благодаря воздушным пузырям и длинным ветвям основная часть слоевища макроцистиса плавает у поверхности моря. Его добывают во всем мире и перерабатывают для получения альгинатов и других химических продуктов.
Нереоцистис (Nereocystis) представлен длинным стволом (черешком) до 15-25 м, который постепенно расширяется кверху, несет на конце пузырь диаметром 12-20 см, от которого отходят узкие пластинки (до 9 м).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 |
