


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
3. Диплоидный. На диплоидном вегетативном талломе после мейоза формируются гаметы (т. е. это гаметический мейоз). Зигота прорастает в новый диплоидный таллом без периода покоя. Гаплоидны только гаметы (диатомовые водоросли, некоторые бурые водоросли).
4. Гаплоидно-диплоидный. Чередование гаплоидных и диплоидных талломов. На гаплоидном талломе формируются споры и гаметы. Первые прорастают в такой же таллом, вторые, сливаясь дают начало диплоидному таллому. На нем формируются диплоидные споры. Они прорастают в такой же таллом. Кроме того формируются гаплоидные споры после мейоза (спорическая редукция), прорастающие в гаплоидный таллом.
Гаплоидный таллом, формирующий гаметы, называют гаметофитом, диплоидный таллом, образующий споры – спорофит.
В зависимости от от морфологии гаметофита и спорофита может быть несколько типов гаплоидно-диплоидного цикла:
4.1. Изомофный. Гаметофит и спорофит морфологически неразличимы. Наиболее распространен.
4.2.. Гетероморфный. Гаметофит и спорофит морфологически различаются, один из них может быть микроскопическим, другой макроскопическим, либо они оба микроскопические.(бурые водоросли Laminaria, Cutleria; красная водоросль Porphyra).
Смена генераций водорослей позволяет гибко реагировать на изменения условий среды. Гаплоидные особи быстрее размножаются, поэтому для них благоприятны условия, способствующие r-отбору. Диплоидные организмы более устойчивы к неблагоприятным условиям среды, способствующим отбору к-стратегов.
Водоросли благодаря образованию спор и гамет могут наряду с чередованием гаплоидных и диплоидных генераций длительное время существовать только в одной какой-либо фазе. Переход от одной фазы цикла к другой у многих водорослей обусловлен адаптациями к изменившимся условиям среды.
Лекция 4.
Красные водоросли – Rhodophyta
Золотистые водоросли – Chrysophyta
Желто-зеленые водоросли – Xanthophyta
Отдел красные водоросли – Rhodophyta
Красные водоросли, своеобразная группа морских макрофитов, насчитывающая около 4000 видов. Своеобразие заключается, прежде всего, в наборе пигментов. Первая эволюционная линия водорослей - красные водоросли (багрянки). Это почти исключительно морские формы, около 3-5% видов обитают в пресных водоемах, единичные представители – на почве. Это естественная, единая древняя группа.
Талломы одноклеточные, в том числе колониальные и многоклеточные. Подавляющее большинство – многоклеточные крупные формы с талломами сложной анатомической и морфологической структуры. Тип структуры таллома – коккоидный, нитчатый, разнонитчатый, пластинчатый. В основе строения многих сложно организованных форм лежит разнонитчатая или гетеротрихальная структура. Дальнейшее усложнение строения однорядного нитчатого таллома – возникновение большого числа обильно разветвленных веточек ограниченного роста на нитях неограниченного роста. Талломы багрянок с большим количеством веточек ограниченного роста образуют значительное количество слизи, которая скрепляет все разветвления, таллом в этом случае приобретает вид слизистого шнура.
При дальнейшем усложнении строения талломов веточки ограниченного роста образуют кору. Кроме того, могут развиваться ризоидные нити или гифы вдоль осевых нитей.
По особенностям анатомического строения различают два основных типа таллома – одноосевой и многоосевой или фонтанный. Принцип построения объемного тела из нитей реализовали высокоорганизованные водоросли, а также грибы, в том числе лишайники.
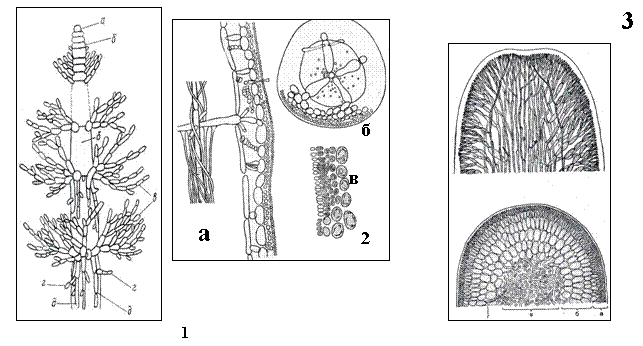

1 - Одноосевой тип таллома (Sirodotia, Batrachospermum): а – инициальная клетка, б – клетки центральной оси, в – ветви ограниченного роста, г – образование междоузлий д – нити коры на междоузлиях; 2 – одноосевой тип таллома (Lemanea): а – часть продольного разреза, б – поперечный разрез, в – разрез через кору с антеридиями; 3 – многоосевой тип таллома (Furcellaria) в продольном и поперечном разрезе.
Клетка красных водорослей одета оболочкой, пектиновые и гемицеллюлозные компоненты которой сильно набухают и часто сливаются в общую слизь мягкой или хрящеватой консистенции. Нередко в стенках откладывается известь. При делении клеток у большинства красных водорослей в стенках остаются неутолщенные места - первичные поры, у более примитивных их нет.
Своеобразие заключается, прежде всего, в наборе пигментов, наряду с хлорофиллами а и d, каротинами и ксантофиллами багрянки содержат специфические - красный фикоэритрин и синий фикоцианин. Различное сочетание этих пигментов определяет окраску водорослей от ярко-красной до голубовато-зеленой и желтой.
Хлоропласты обладают оболочкой из двух мембран. Фикобилины локализованы в фикобилисомах, расположенных на поверхности тилакоидов. В «Биологическом энциклопедическом словаре и «Жизни растений» ошибочно указано, что К. В. содержат хлорофилл b, то не удивляйтесь, - это ошибка. Их хлоропласты произошли, по-видимому, от симбиотических цианобактерий. Хлоропласты с пиреноидами или без них. Тилакоиды расположены в хлоропластах одиночно.
Запасной продукт – багрянковый крахмал. Его зерна откладываются вне хлоропластов – в цитоплазме. Жгутиков нет даже у гамет.
Размножение. Для жизненного цикла К. В. характерно отсутствие жгутиковых стадий. Вегетативное размножение осуществляется делением клеток, фрагментацией слоевищ. Бесполое размножение осуществляется посредством спор. Моноспорангии, биспорангии, тетраспорангии и полиспорангии. Моноспоры, биспоры, тетраспоры и полиспоры.
Смена разных форм размножения, чередование бесполого и полового размножения. Смена гаплоидной и диплоидной фаз. Жизненный цикл или цикл развития. Спорофиты и гаметофиты.
Тип полового процесса красных водорослей – оогамия. Карпогон, трихогина, антеридии (сперматангии), спермации, сорусы, процесс оплодотворения, зигота, карпоспоры.
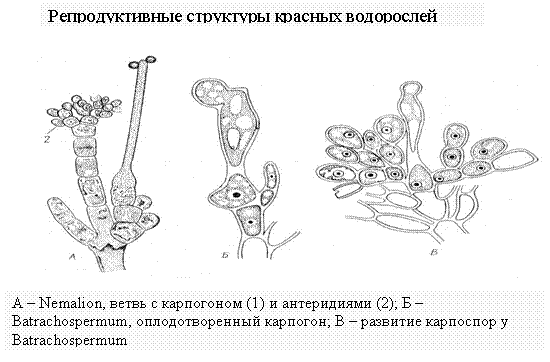

Женский орган - карпогон, состоит из расширенной базальной части - брюшка и отростка - трихогины. Карпогон обычно развивается на особой короткой карпогониальной ветви. Антеридии - мелкие, бесцветные клетки, внутри которых созревают голые, лишенные жгутиков, спермации. Спермации пассивно переносятся током воды, прилипают к трихогине. В месте контакта стенки растворяются, ядро спермация перемещается до яйцеклетки, где происходит слияние ядер. Дальнейшее развитие приводит к образованию карпоспор. Детали этого развития имеют важное систематическое значение. У одних красных водорослей содержимое зиготы (оплодотворенного карпогона) непосредственно делится с образованием неподвижных голых карпоспор, у других из оплодотворен-ного карпогона вырастают ветвящиеся нити - гонимобласты, клетки которых превращаются в карпоспорангии, производящие по одной карпоспоре. У большинства же красных водорослей гонимобласты развиваются не непосредственно из брюшка оплодотворенного карпогона, а из особых вспомогательных - ауксилярных клеток. Последние могут быть удалены от карпогона или же располагаются на талломе в непосредственной близости от него. В тех случаях, когда ауксилярные клетки удалены от карпогона, из его брюшка после оплодотворения вырастают соединительные, или ообластемные, нити. Их возникновению предшествует митотическое деление копуляционного ядра, и клетки ообластенных нитей содержат дипло-идные ядра. Ообластемные нити подрастают к ауксилярным клеткам, в точке контакта оболочки растворяются и между клеткой ообластемной нити и ауксилярной клеткой устанавливается сообщение. Это слияние клеток не сопровождается слиянием их ядер (диплоидного ядра клетки ообластемной нити и гаплоидного ядра ауксилярной). Однако слияние с ауксилярной клеткой стимулирует деление диплоидного ядра клетки ообластемной нити и развитие гонимобластов, клетки которых содержат диплоидные ядра и продуцируют диплоидные карпоспоры. Гонимобласты, развивающиеся карпоспоры, обычно рассматриваются как особая генерация - карпоспорофит. У наиболее высокоорганизованных красных водорослей ауксилярные клетки развиваются только после оплодотворения карпогона и в непосредственной близости от него. Ауксилярная клетка сливается с брюшком оплодотворенного карпогона, после чего из нее развиваются гонимобласты с карпоспорами. Карпоспорангии часто располагаются тесными группами - цистокарпиями, которые одеты псевдопаренхимной оболочкой.
Для высокоорганизованных красных водорослей характерны очень сложные циклы развития, при которых разные стадии, в частности гаметофит и спорофит, могут выглядеть совершенно различно. Ранее они были описаны как разные неродственные рода. Многообразие циклов развития лежит в основе систематики этой группы.
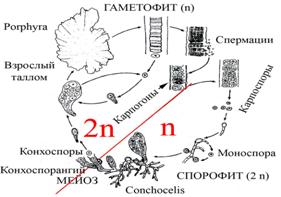
Два варианта жизненных циклов красных водорослей на примере родов Porphyra и Polysiphonia, которые являются представителями двух классов красных водорослей.
Гаплоидный набор хромосом на схемах обозначен как n, диплоидный – как 2 n.
Характеристика жизненных циклов Porphyra и Polysiphonia.
Схема жизненного цикла Porphyra



Красные водоросли - типичные морские растения, но отдельные представители живут в прудах и реках, лужах и озерах, есть и наземные организмы. В морях красные водоросли поселяются на твердых неподвижных грунтах (скалы, рифы, валуны, каменистые россыпи). Среди красных водорослей распространены явления эпифитизма и паразитизма. При подходящем грунте и хорошей прозрачности воды они достигают глубины 100-200 м. Гораздо чаще заросли красных водорослей кончаются на глубине 20-40 м. Растут во всех морях Мирового океана от тропиков до полюсов. Наиболее разнообразно они представлены в тропиках. Существует теория так называемой хроматической адаптации, по которой проникновение водорослей на те или иные глубины связано с количеством света, проходящего через толщу воды. Как известно, глубже всего проникают лучи из зеленой и синей частей спектра. Красные пигменты багрянок позволяют им фотосинтезировать в синих лучах, и поэтому, они проникают на глубины, недоступные для других водорослей. В опытах было установлено, что при слабом освещении они участвуют в усиленном поглощении света. Проникновение красных водорослей на значительные глубины объясняется их способностью усваивать малые количества света. В целом, багрянки - теневыносливые организмы. Для защиты от сильного света у багрянок, живущих на небольших глубинах, служат особые иридирующие тельца (мелкие зернышки протеиновой природы в вакуолях). Иридирующие тельца способны рассеивать и отражать падающие на них солнечные лучи. Водоросли, обладающие иридирующими тельцами, имеют обычно в падающем свете голубоватостальной блеск.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 |
