

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
GPP и, следовательно, скорость фотосинтеза при одном и том же уровне ФАР была выше в утренние часы относительно постполуденных (рис. 7). Эта полуденная депрессия фотосинтеза была ярко выражена не только в месяцы сухого сезона, но и в течение влажного периода. Например, в марте в утренние часы усреднённая GPP при ФАР 1000 мкмоль/м2с составляла 19 мкмоль/м2с, в то время как в постополуденные время снижалась на 34% и составляла только 12,5 мкмоль/м2с при том же уровне ФАР. В июле в постполуденное время GPP снижалась в среднем на 36%, значения GPP составили 27,5 мкмоль/м2с и 17,5 мкмоль/м2с до и после полудня соответственно. Таким образом, как в сухой, так и во влажный сезон картина снижения GPP в постполуденный период времени была сходна.
2. Температура воздуха. Исследована зависимость GPP от температуры воздуха на высоте 30 м. GPP достоверно коррелирует с температурой воздуха как в сухой, так и во влажный сезоны (R2 = 0,22 и 0,27 соответственно), особенно сильная зависимость наблюдалась в утренние, предполуденные часы, когда R2 может достигать 0,5. При этом с ростом температуры воздуха растёт и интенсивность фотосинтеза, но при определённых значениях температуры воздуха наблюдается ингибирование фотосинтетической активности. Температурное ингибирование при 30 °С отмечено в течение всего года.
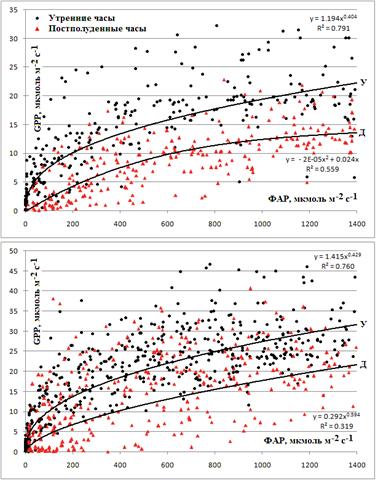
Рис. 7. Полуденная депрессия фотосинтеза в марте (сверху) и июле (снизу). Показана зависимость GPP (в мкмоль/м2с) от ФАР (мкмоль/м2с). Регрессионный анализ проведён отдельно для утреннего периода (до 11 ч местного времени, У) и для постполуденного периода (после 13 ч местного времени, Д)
3. Дефицит водяных паров в воздухе (VPD). Анализ зависимости GPP от дефицита водяных паров в воздухе показал явное ингибирование фотосинтетической активности при высоких значениях VPD как в сухой (март), так и во влажный (июль) сезоны. В марте ингибирование начиналось при VPD 12 гПа, в июле – при VPD 8-10 гПа.
3.2.3. Сезонная динамика GPP
Изменение GPP по месяцам показано на рис. 8.
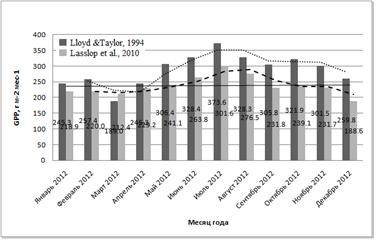
Рис. 8. Суммарная первичная валовая продукции углерода (в г/м2 в месяц) в разные месяцы года, определённая с использованием двух различных способов разделения потоков
Изменение валовой продукции в течение года было небольшим – от 219 г/м2 за месяц в марте до 374 г/м2 в месяц в июле. Наблюдается тенденция некоторого увеличения GPP в течение влажных месяцев, после чего, с резким падением количества осадков, величина продукции постепенно снижается, достигая типичных для сухого сезона относительно небольших значений.
Роль внешних факторов.
1. ФАР. Выявлена небольшая (R2=0,18) достоверная зависимость суточных сумм GPP от суточных сумм ФАР в сезонной динамике (рис. 9). GPP выходит на плато уже при относительно небольшом ФАР (~25 моль/м2сутки). Подобные суммы достигаются почти ежедневно даже в течение влажного сезона (рис. 10).
2. Влажность почвы на глубине 50 см. Не удалось обнаружить значимой корреляции между GPP и влажностью почвы на глубине 50 см.
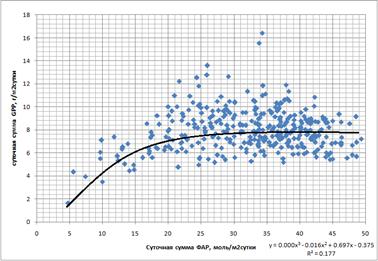
Рис. 9. Обобщённая зависимость суточных сумм GPP (гС/м2сутки) от суточных сумм ФАР (моль/м2сутки)
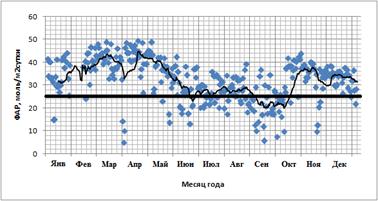
Рис. 10. Изменение суточной суммы ФАР (моль/м2сутки) в течение 2012 года. Жирной линией отмечен уровень насыщения ФАР (25 моль/м2сутки).
Глава 4. Обсуждение результатов
4.1. Таксационные описания
Проведённые таксационные описания выявили различия в биомассе между разными типами лесных сообществ. Полученные данные по биомассе отдельных типов лесных сообществ согласуются с данными литературы по аналогичным сообществам в других экваториальных странах (FAO Forestry paper-134, 1997).
Анализ картографического материала показал, что на территории, где расположен измерительный комплекс для пульсационных исследований, преобладает естественно восстанавливающийся лес, но значительную площадь (10-20%) занимают и два других изученных сообщества (лагерстремовый лес и редколесье). Данное распределение можно считать типичным только для северо-восточной равнинной части парка, но не для всего Кат Тьена в целом (где в значительной мере присутствуют территории, покрытые бамбуком, и другие типы леса).
На основании таксационных описаний можно сделать вывод о том, что в значительной мере леса парка обладают неполным потенциалом биомассы. При этом наибольший вклад в депонирование углерода можно ожидать в результате развития естественно восстанавливающихся древостоев.
Возможно, значительные части «редколесья» образовались в результате мощной деградации в прошлом, однако одной из важных причин являются и природные условия соответствующих территорий (приуроченность к более сырым, заболоченны участкам). Из-за этого потенциал развития у редколесья не должен быть большим, и значительный вклад в будущее депонирование углерода в лесных экосистемах парка этот тип леса оказать не сможет.
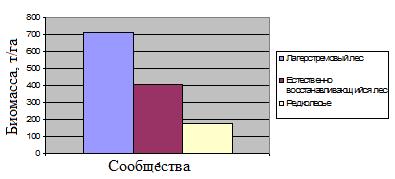
Рис. 11. Общее распределение биомассы в изученных сообществах (т/га).
4.2. Суточная динамика GPP
Максимальные средние суточные значения GPP в течение всех месяцев были сравнимы (~20-35 мкмоль/м2с по методу разделения потоков Lloyd and Taylor, ~15-35 мкмоль/м2с по методу разделения потоков Lasslop. Достижение точки максимума и диапазон области оптимума значений GPP включает время от 9 до 14 ч (по методу Lloyd-Taylor) и от 8 до 13 ч (по методу Lasslop). Сравнение двух методов показало, что при использования метода разделения по Lasslop оптимум сдвигается на более раннее время суток. Следует отметить, что диапазон оптимума по времени сходен и отмечается в обоих вариантах обработки полученных результатов. Коч (Koch et al., 1994), работавший в Панаме и изучавший суточную динамику фотосинтеза деревьев полога, обнаружил, что максимум фотосинтеза наблюдался в 11 часов местного времени, что также смещено относительно астрономического полудня. Сходные с нашими данные по суточной динамике GPP были получены и для других тропических лесов (Grace, 1995; Bonal et al., 2008; Desai et al., 2008; Goulden et al., 2004).
В марте кривая фотосинтеза выходит на уровень насыщения при значениях ФАР 900-1000 мкмоль/м2с, что составляет около 50% от возможных максимальных значений. Диапазон варьирования значений GPP при этом значении ФАР составляет от 12 до 35 мкмоль/м2с (в среднем около 20 мкмоль/м2с). При данном уровне ФАР присутствуют единичные максимальные значения (30-35 мкмоль/м2с). В июле насыщение начинается несколько позже, оно хорошо выражено в диапазоне ФАР 1000-1200 мкмоль/м2с при средних значениях GPP 25-30 мкмоль/м2с (варьирование от 20 до 45 мкмоль/м2с). Очень высокие значения GPP (более 40 мкмоль/м2с) также, как и в марте, являются редкими и заметно выделяются из основного массива данных. Наши данные согласуются с литературными. Например, (Goulden еt al., 2004), отметил начало насыщения при ФАР 500-1000 мкмоль/м2с для зрелых тропических лесов Амазонии. Сходные результаты получены и другими авторами (Malhi et al. 1998, Fan et al. 1990).
Наличие полуденной депрессии фотосинтеза показано для множества экосистем (Larcher, 1995), включая и тропические леса (Koch et al. 1994, Malhi et al. 1998, Zotz and Winter 1996). Исследователи объясняют это явление множеством факторов, включающих устьичный ответ на уменьшение давления водяных паров в воздухе, низкий водный потенциал листа, влияние на фотосинтез повышенной температуры воздуха, накоплением метаболитов и крахмала, внутренние циркадные ритмы и др. Влияние этих факторов является комплексным. Несмотря на это, наши данные позволяют рассмотреть подробнее влияние некоторых факторов на особенности суточного хода GPP в муссонном тропическом лесу южного Вьетнама.
Вероятно, одним из определяющих факторов в формировании полуденной депрессии фотосинтеза является рост дефицита водяных паров в воздухе (VPD). В марте максимальные значения GPP отмечаются при VPD на уровне 12 гПа, что соответствует оптимальным средним значениям GPP – около 17 мкмоль/м2с, после чего отмечается ингибирование фотосинтетической активности. В июле в широком диапазоне водного дефицита (от 1 до 12 гПа) изменения в величине GPP небольшие, но максимальные значения продукции отмечаются уже в области 7-8 гПа, которым соответствует средний оптимальный уровень GPP 23 мкмоль/м2с. Более низкий водный дефицит во влажный сезон (июль) способствует и более поддержанию значений GPP на более высоком уровне. Таким образом, во влажный сезон наблюдается меньшая зависимость GPP от этого фактора. К такому же выводу пришли и Koch с соавторами (1994), которые изучали фотосинтез и проводимость листа деревьев полога в лесах Панамы. Они обнаружили, что фотосинтез главным образом определяется проводимостью листа, которая в первую очередь зависит от VPD (Koch et al., 1994).
Высокая температура воздуха также может являться фактором, ограничивающим интенсивность фотосинтеза. При температурах воздуха выше 30 °С в течение всего года GPP начинает падать. Эта закономерность в большей степени выражена в июле, когда падение после насыщения более резкое. Вероятно, падение интенсивности фотосинтеза в данном случае связано не с повышением температуры воздуха, а с увеличением VPD. Влияние температуры на скорость фотосинтеза многократно описывалась различными исследователями, которые пришли к выводам, что температура до 30-35 °С ещё не ограничивает интенсивность фотосинтеза. Например, Lloyd and Farquhar (2008) показали, что температура 30 °С сама по себе не ограничивает скорость фотосинтеза тропических деревьев Амазонского бассейна.
4.3. Сезонная динамика GPP. В сезонном аспекте GPP варьирует в пределах от 220 гС/м2 в месяц в марте до 300-375 гС/м2 в месяц в июле-августе. При этом месячные суммы продукции в сухие месяцы несколько меньше, чем суммы, полученные во влажные месяцы (Рис. 8).
Стоит отметить, что максимальные месячные суммы GPP леса национального парка Кат Тьен в течение влажного сезона сравнимы с месячными суммами GPP широколиственных лесов в период пика вегетации. Например, в лиственничном лесу Томакомай (Tomakomai, Япония) самым продуктивным месяцем является июнь с GPP ~420 гС/м2 месяц (Saigusa et al., 2008), что выше, чем максимальная месячная продуктивность муссонного тропического леса национального парка Кат Тьен. Таким образом, общие высокие годовые суммы GPP в тропиках достигаются преимущественно не за счёт собственной высокой продуктивности, а за счёт того, что период вегетации длится в течение всего года, без периодов покоя. Это имеет место в том числе в сезонно-влажных тропиках, где наблюдается лишь относительно небольшое снижение GPP в течение засушливого периода (Bonal et al., 2008).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
