

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Число семян в почвенном запасе, хотя этот показатель и очень важен, при построении возрастного спектра ценопопуляции обычно не учитывается, так как подсчет их очень трудоемок и получить статистически надежные величины практически невозможно.
Если в возрастном спектре ценопопуляции в момент ее наблюдения представлены только семена или молодые особи, ее называют инвазионной. Такая ценопопуляция не способна к самоподдержанию, и существование ее зависит от поступления зачатков извне. Часто это молодая ценопопуляция, только что внедрившаяся в биоценоз. Если ценопопуляция представлена всеми или почти всеми возрастными группами (некоторые возрастные состояния у конкретных видов могут быть не выражены, например, имматурное, субсенильное, ювенильное), то она называется нормальной. Такая популяция независима и способна к самоподдержанию семенным или вегетативным путем. В ней могут преобладать те или иные возрастные группы. В связи с этим различают молодые, средневозрастные и старые нормальные ценопопуляции.
Нормальная ценопопуляция, состоящая из особей всех возрастных групп, называется полночленной, а если особи каких-либо возрастных состояний отсутствуют (в неблагоприятные годы временно могут выпадать отдельные возрастные группы), то популяция называется нормальной неполночленной.
Регрессивная ценопопуляция представлена только сенильными и субсенильными или также генеративными, но старыми, не образующими всхожих семян. Такая ценопопуляция не способна к самоподдержанию и зависит от заноса зачатков извне.
Инвазионная ценопопуляция может перейти в нормальную, а нормальная – в регрессивную.
Возрастная структура ценопопуляции во многом определяется биологическими особенностями вида: периодичностью плодоношения, числом продуцируемых семян и вегетативных зачатков, способностью вегетативных зачатков к омоложению, скоростью перехода особей из одного возрастного состояния в другое, способностью образовывать клоны и др. Типичный возрастной спектр называется базовым (рис. 99). Проявление всех этих биологических особенностей, в свою очередь, зависит от условий внешней среды. Меняется и ход онтогенеза, который может протекать у одного вида во многих вариантах (поливариантность онтогенеза), что сказывается на структуре возрастного спектра ценопопуляции (рис. 100).
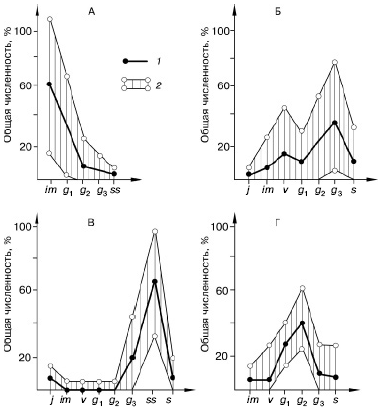
Рис. 99. Базовый тип спектра ценонопуляций (по , 1976) А – бурачок ленский; Б – анабазис безлистный; В – овсяница луговая; Г – типчак.
1 – базовый спектр; 2 – пределы изменения базового спектра
Разные размеры растений отражают различную жизненность особей в пределах каждой возрастной группы. Жизненность особи проявляется в мощности ее вегетативных и генеративных органов, что соответствует количеству накопленной энергии, и в устойчивости к неблагоприятным воздействиям, что определяется способностью к регенерации. Жизненность каждой особи меняется в онтогенезе по одновершинной кривой, возрастая на восходящей ветви онтогенеза и уменьшаясь на нисходящей. У многих видов особи одного возрастного состояния в одной ценопопуляции могут иметь разную жизненность. Эта дифференциация особей по жизненности может вызываться разнокачественностью семян, разными сроками их прорастания, микроусловиями среды, воздействием животных и человека, конкурентными отношениями. Высокая жизненность может сохраняться до смерти особи во всех возрастных состояниях или снизиться в ходе онтогенеза. Растения высокого уровня жизненности нередко проходят все возрастные состояния ускоренными темпами. В ценопопуляциях часто преобладают растения среднего уровня жизненности. Одни из них проходят онтогенез полностью, а другие пропускают часть возрастных состояний, переходя перед отмиранием на более низкий уровень жизненности. Растения низшего уровня жизненности имеют сокращенный онтогенез и часто переходят в сенильное состояние, едва приступив к цветению.
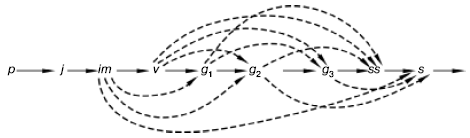
Рис. 100. Варианты развития ежи сборной в разных экологических условиях (по , 1985). Латинскими буквами обозначены возрастные состояния растений, а пунктирными линиями – их возможная последовательность
Особи одной ценопопуляции могут развиваться и переходить из одного возрастного состояния в другое с разной скоростью. По сравнению с нормальным развитием, когда возрастные состояния сменяют друг друга в обычной последовательности, может наблюдаться ускорение или задержка в развитии, выпадение отдельных возрастных состояний или целых периодов, наступление вторичного покоя, часть особей может омолаживаться или отмирать. Многие луговые, лесные, степные виды при выращивании их в питомниках или посевах, т. е. на лучшем агротехническом фоне, сокращают свой онтогенез, например овсяница луговая и ежа сборная – с 20–25 до 4 лет, горицвет весенний – со 100 до 10–15 лет, жабрица порезниковая – с 10–18 до 2 лет. У других растений при улучшении условий онтогенез может удлиняться, как, например, у тмина обыкновенного.
В засушливые годы и при усилении выпаса у степного вида овсеца Шелля отдельные возрастные состояния выпадают. Например, взрослые вегетативные особи могут сразу пополнять группу субсенильных, реже – старых генеративных. Клубне-луковичные растения безвременника великолепного в центральных частях компактных клонов, где условия менее благоприятные (хуже освещение, увлажнение, минеральное питание, проявляется токсическое действие отмерших остатков), быстрее переходят в сенильное состояние, чем периферические особи. У свербиги восточной при усиленной пастбищной нагрузке, когда повреждаются почки возобновления, молодые и зрелые генеративные особи могут иметь перерывы в цветении, тем самым как бы омолаживаясь и продлевая свой онтогенез.
У ежи сборной в разных условиях реализуется от 1–2 до 35 путей онтогенеза, а у подорожника большого от 2–4 до 100. Возможность менять путь онтогенеза обеспечивает адаптацию к меняющимся условиям среды и расширяет экологическую нишу вида.
У двух видов степного овсеца – Шелля и опушенного – в Пензенской области четко прослежено циклическое изменение возрастных спектров в многолетней динамике. В засушливые годы популяции овсецов стареют, а во влажные – молодеют. Флюктуации возрастного спектра ценопопуляций вслед за погодными условиями особенно характерны для растений поемных лугов.
Возрастной спектр может варьировать не только вследствие внешних условий, но и в зависимости от реактивности и устойчивости самих видов. К выпасу у растений разная сопротивляемость: у одних выпас вызывает омолаживание, так как растения отмирают, не достигнув старости (например, у полыни равнинной), у других – способствует старению ценопопуляции изза снижения возобновления (например, у степного вида жабрицы Ледебура).
У некоторых видов на всем протяжении ареала в широком диапазоне условий нормальные ценопопуляции сохраняют основные черты возрастной структуры (ясень обыкновенный, типчак, овсяница луговая и др.). Такой возрастной спектр зависит преимущественно от биологических свойств вида. В нем сохраняются прежде всего соотношения во взрослой, наиболее стабильной части. Число вновь возникающих и отмирающих особей в каждой возрастной группе сбалансировано, и общий спектр оказывается постоянным до значительных изменений условий существования. Такие базовые спектры чаще всего имеют ценопопуляции видов-эдификаторов в устойчивых сообществах. Им противопоставляются ценопопуляции, относительно быстро меняющие возрастной спектр вследствие не установившихся отношений с окружающей средой.
Чем крупнее особь, тем значительнее сфера и степень ее воздействия на среду и на соседние растения («фитогенное поле», по ). Если в возрастном спектре ценопопуляции преобладают особи взрослые вегетативные, молодые и средневозрастные генеративные, то вся популяция в целом будет занимать более прочное положение среди других.
Таким образом, не только численность, но и возрастной спектр ценопопуляции отражает состояние и приспособленность ее к меняющимся условиям внешней среды и определяет позиции вида в биоценозе.
Возрастная структура популяций у животных. В зависимости от особенностей размножения члены популяции могут принадлежать к одной генерации или к разным. В первом случае все особи близки по возрасту и примерно одновременно проходят очередные этапы жизненного цикла. Примером может служить размножение многих видов нестадных саранчовых. Весной из яиц, перезимовавших в кубышках, отложенных в землю, появляются личинки первого возраста. Отрождение личинок несколько растягивается под влиянием микроклиматических и иных условий, но в целом протекает довольно дружно. В это время популяция состоит только из молодых насекомых. Через 2–3 недели в связи с неравномерностью развития отдельных особей в ней могут одновременно встречаться личинки смежных возрастов, но постепенно вся популяция переходит в имагинальное состояние и к концу лета состоит только из взрослых половозрелых форм. К зиме, отложив яйца, они погибают. Такова же возрастная структура популяций у дубовой листовертки, слизней рода Deroceras и других видов с однолетним циклом развития, размножающихся один раз в жизни. Сроки размножения и прохождения отдельных возрастных стадий обычно приурочены к определенному сезону года. Численность таких популяций, как правило, неустойчива: сильные отклонения условий от оптимума на любой стадии жизненного цикла действуют сразу на всю популяцию, вызывая значительную смертность.
Виды с одновременным существованием различных генераций можно разделить на две группы: размножающиеся один раз в жизни и размножающиеся многократно.
У майских жуков, например, самки вскоре после откладки яиц весной погибают. Личинки развиваются в почве и окукливаются на четвертый год жизни. Одновременно в популяции присутствуют представители четырех генераций, каждая из которых появляется через год после предыдущей. Ежегодно завершает свой жизненный цикл одна генерация и появляется новая. Возрастные группы в такой популяции разделяются четким интервалом. Соотношение их по численности зависит от того, насколько благоприятными оказались условия при появлении и развитии очередного поколения. Например, генерация может оказаться малочисленной, если поздние заморозки погубят часть яиц или холодная дождливая погода помешает лету и копуляции жуков.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 |
