

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На границах своего временного ареала перелетная саранча не может размножаться, и эти очаги вскоре затухают. Стаи или погибают, или, постепенно разреживаясь, начинают откочевку к зоне постоянных очагов. В изреженных популяциях вновь происходит переход к одиночной фазе, через 2–3 промежуточных поколения. Таким образом, разлет стай саранчи не обеспечивает формирования новых постоянных популяций, а практически служит лишь механизмом снятия перенаселенности в местах, благоприятных для размножения. При этом гибнет гигантское количество насекомых. Стаи, несущие на своем пути неисчислимые бедствия, сами оказываются обреченными.

Рис. 132. Нашествие перелетной саранчи
Явление фазовости обнаружено не только у стадных саранчовых, но и у других беспозвоночных. У тлей увеличение плотности населения вызывает появление крылатой фазы и разлет насекомых с образованием новых поселений. Обычно тли дают несколько поколений бескрылых самок, но в условиях постоянного перенаселения крылатые самки развиваются в каждом поколении. У ряда амеб химические изменения состава водной среды, вызванные переуплотнением популяции, стимулируют переход в подвижную жгутиковую стадию. В результате происходит быстрое рассредоточение особей в пространстве.
Территориальное поведение животных, выработавшееся в ходе эволюции как система инстинктов, – наиболее эффективный механизм сдерживания роста численности популяции на данной площади. Мечение и охрана участков, не допускающие размножения на них «чужих» особей, приводят к рациональному использованию территории. Избыточная часть популяции при этом не размножается или вынуждена выселяться за пределы занятого пространства. Это же относится и к выведенному потомству, среди которого лишь некоторая часть вследствие естественной смертности взрослых занимает освобождающиеся участки.
Выселения как ответная реакция на растущую плотность популяции свойственны многим видам птиц и млекопитающих. Кроме обычной расселительной дисперсии молодняка, для ряда видов с резкими колебаниями численности характерны массовые перемещения – нашествия. Они возникают нерегулярно, лишь в годы вспышек размножения, и не имеют постоянного направления. Такие нашествия описаны, например, у тундровых леммингов, белок Сибири и Северной Америки и др. При нашествиях часть особей остается на месте, а среди эмигрантов преобладают молодые.
Повышение плотности популяции может сопровождаться такими изменениями в физиологии особей, которые ведут к снижению рождаемости и увеличению смертности. У млекопитающих известно явление стресса, которое впервые было описано в 1936 г. физиологом Г. Селье для человека. В ответ на отрицательное воздействие каких-либо факторов в организме возникают реакции двух типов: 1) специфические, зависящие от природы повреждающего агента (например, усиление теплопродукции при действии холода), и 2) неспецифическая реакция напряжения как общее усилие организма приспособиться к изменившимся условиям. Эта общая реакция складывается из ряда физиологических и морфологических изменений, которые постепенно развертываются как единый процесс. Реакция напряжения, или стресс, возникает в ответ на любые отрицательные воздействия среды, в том числе и на отклонение плотности популяции от оптимума.
Большую роль в развитии стресса играют сигналы коры головного мозга, меняющие активность гипоталамуса – центрального звена вегетативной нервной системы. В свою очередь, деятельность гипоталамуса вызывает изменения в функционировании гипофизарно-надпочечниковой гормональной системы. В состоянии стресса у животных сильно увеличивается кора надпочечников и повышается концентрация кортикостероидных гормонов, выделяемых этим органом, а также происходит целый ряд других изменений в гормональном равновесии организма. У самок в популяции учащаются нарушения овуляции, резорбция эмбрионов, рано прекращается лактация, угасают инстинкты заботы о потомстве и т. п., уменьшается число выводков и количество в них молодых. В конечном счете все это приводит к торможению роста популяции. У мышевидных грызунов, содержащихся в клетках одинаковых размеров, четко проявляется обратно пропорциональная зависимость между численностью зверьков в клетке и массой их репродуктивных органов. В состоянии стресса у животных даже при достаточном снабжении кормом понижается устойчивость к вредным воздействиям среды, что увеличивает смертность.
От плотности населения зависит в первую очередь поведение животных. У многих видов в условиях скученности повышается уровень агрессивности, меняется реакция на особей противоположного пола, молодняк и т. д.
Стресс-реакция как механизм, регулирующий рождаемость, особенно отчетливо проявляется у животных с хорошо выраженной системой иерархического подчинения в группах.
Реакция напряжения свойственна подчиненным животным; у них также тормозится воспроизводительная функция. Доминирующие особи стресс-реакции не проявляют. В переуплотненных популяциях стресс распространяется на большую часть особей и, по-видимому, тормозит размножение.
Стрессовое состояние не вызывает необратимых изменений в половой системе, а приводит лишь к временному гормональному блокированию ее функции. После устранения перенаселенности способность к размножению может восстановиться в короткие сроки.
Закономерности стресса, вызываемого перенаселением, изучают в основном на лабораторных животных. Однако многочисленные факты, зарегистрированные в природных популяциях, позволяют предполагать, что в естественных условиях стресс играет немалую роль в регуляции численности и структуры популяций и поведения млекопитающих (рис. 133).
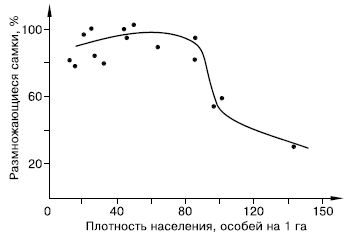
Рис. 133. Зависимость интенсивности размножения от плотности населения в популяции малого суслика (по , 1995)
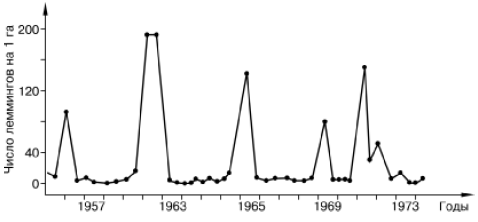
Рис. 134. Динамика численности сибирского лемминга на Аляске (по Bunnel et all., 1975)
Например, в динамике популяций ряда тундровых леммингов зарегистрированы правильные циклы трех-четырехлетней периодичности с амплитудой колебания до 600 крат (рис. 134). Фаза пика в таких колебаниях ограничена обычно одним сезоном, затем следуют резкий спад, фаза депрессии численности и последующего нарастания. При нарастании численности регистрируется увеличение плодовитости, повышение скорости созревания молодняка, усложнение возрастной структуры популяции, снижение смертности новорожденных. В период пика отмечается резкое снижение воспроизводства и одновременно увеличивается смертность во всех возрастных группах. В яичниках самок всех возрастов обнаруживаются серьезные деструктивные изменения, наблюдается массовая гибель фолликулов на ранних стадиях развития. Год-два после спада численности общая интенсивность размножения остается средней, а смертность высокой, а затем вновь все показатели размножаемости начинают расти. У зверьков, родившихся при низкой численности популяции, восстанавливается нормальное функционирование яичников.
На разных этапах этого цикла сильно меняется гормональное состояние зверьков, относящихся к разным генерациям. В пик численности отмечается избыточная активность адреналовой и щитовидной желез, что резко тормозит воспроизводительные функции организма. В ходе цикла у сменяющихся поколений меняется не только функционирование отдельных желез, но и всей эндокринной системы животных. Через несколько поколений, в период минимума численности, состояние эндокринной системы нормализуется и обеспечивает восстановление эффективности процесса размножения. Конкретные причины таких гормональных различий у разных поколений связаны с тем, что жизнеспособность и эндокринные особенности организма формируются в эмбриональный период и во многом определяются физиологическим состоянием родителей.
Таким образом, динамику численности леммингов можно представить как авторегулируемый процесс, в котором большую роль играют эндокринные механизмы.
Все рассмотренные выше примеры взаимодействия между членами популяции, от «жестких» форм – прямого уничтожения одной особью другой – до снижения воспроизводительных способностей как условного рефлекса на повышение частоты контактов, представляют собой разные формы ограничения роста популяций. Эти тормозящие механизмы включаются до полного истощения ресурсов среды в ответ на сигналы, свидетельствующие об угрозе перенаселения.
Степень развития механизмов популяционного гомеостаза находится также в тесной связи с тем, насколько влияют на популяцию другие виды: конкуренты, хищники, паразиты. Общая регуляция численности популяций в природных сообществах происходит в результате сложных межвидовых и внутривидовых взаимоотношений.
8.6. Регуляция численности популяций в биоценозах
8.6.1. Модификация и регуляция популяций
Современная теория рассматривает динамику численности популяций как авторегулируемый процесс. Любой популяции организмов в конкретных условиях свойствен определенный средний уровень численности, вокруг которого происходят колебания. Отклонения от этого среднего уровня имеют разный размах, но в норме после каждого отклонения численность популяции начинает изменяться с обратным знаком. Теория динамики численности разрабатывается в основном на примере животных, хотя целый ряд ее положений применим и к растениям, грибам, простейшим и прокариотам.
Выделяют две принципиально разные стороны популяционной динамики: модификацию и регуляцию. Модификация – это случайное отклонение численности, возникающее в результате воздействия самых разнообразных факторов, не связанных с плотностью популяции. Регуляция – это возврат популяции после отклонения к исходному состоянию, совершающийся под влиянием факторов, сила действия которых определяется плотностью популяции.
Модифицирующие факторы, вызывая изменение численности популяций, сами не испытывают влияния этих изменений. Действие их, таким образом, одностороннее. К ним относятся все абиотические влияния среды на организмы, на качество и количество их корма и т. п. Благоприятная погодная обстановка может послужить причиной массовой вспышки размножения вида и перенаселения занимаемой им территории, как, например, в случае стадных саранчовых. Отрицательное воздействие модифицирующих факторов, наоборот, снижает численность популяции иногда до полного ее исчезновения.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 |
