

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Ранг животного в группе определяется столкновениями между особями, которые могут иметь характер прямой борьбы или ритуальных угроз. После установления ранга всех членов группы прямые столкновения между ними прекращаются и порядок поддерживается сигнальным или ритуальным поведением.
Иерархически организованному стаду свойствен закономерный порядок перемещения, определенная организация при защите, расположении на местах отдыха и т. п. Так, при передвижении семейного стада зебр впереди всегда идет старая кобыла, за ней следуют сначала самые молодые, а затем старшие жеребята, за которыми двигаются взрослые зебры. Шествие завершает жеребец-вожак. В стаде павианов в центре, в наибольшей безопасности, находятся самки с детенышами или готовящиеся к размножению, по краям – вожаки, молодые самцы и неразмножающиеся самки (рис. 119). Впереди и позади стада шествуют крупные самцы, готовые отразить нападение. Описаны случаи, когда при преследовании стада хищниками самец-доминант возвращался за отставшим детенышем, несмотря на грозившую ему большую опасность.


Рис. 119. Походный порядок стада павианов (по , 1972):
вверху – в пути; внизу – при нападении леопарда.
С – самки; М – молодняк; В – вожаки; Н – самцы низшего ранга
Вожаки-доминанты выполняют разнообразные общественные функции. В табуне лошадей, например, вожак управляет движением, уводит косяк от опасности, защищает от хищников, прекращает драки, проявляет заботу о жеребятах и больных животных и т. п. В сложной обстановке вожаки ряда видов (лошади Пржевальского, павианы) могут проводить разведку, отделяясь от стада, а затем возвращаясь к нему и определяя безопасное направление движения.
Конфликты в группах, имеющих вожака, обычно решаются с его участием или в его присутствии, у разных видов с разной степенью агрессивности. В стадах павианов доминанты могут прибегать к преследованию и укусам, тогда как у горилл вожак нередко восстанавливает порядок лишь взглядом или движением головы.
Биологический смысл иерархической системы доминирования-подчинения заключается в создании согласованного поведения группы, выгодного для всех ее членов. После «расстановки сил» животные не тратят лишней энергии на индивидуальные конфликты, а группа в целом получает преимущества, подчиняясь наиболее сильным и опытным индивидуумам (рис. 120). Согласованность поведения имеет большое значение в обеспечении защиты от хищников, предупреждении опасности, выращивании молодняка, миграциях и т. п. (рис. 121). В сложных условиях, например при голодовках, гибнут в первую очередь более слабые, подчиненные особи, но под защитой группы они все же имеют больше шансов выжить, чем в одиночку.
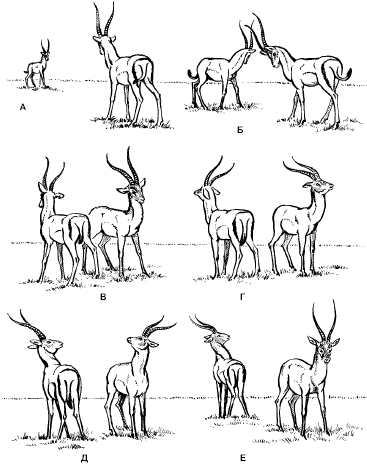
Рис. 120. Встреча двух самцов газели Гранта (по Р. Шовену, 1972):
А-Д – позы, соответствующие разным фазам ритуала противостояния;
Е – самец, повернувший голову, – победитель

Рис. 121. Индивидуальные дистанции и направления осматривания у архаров во время отдыха (по , 1976)
Иерархия в групповых отношениях адаптивна прежде всего потому, что она динамична. В жизни стада идет непрерывная проверка соответствия его структуры меняющимся условиям. Состав группы также не остается постоянным: взрослеет молодняк, появляются новые пришельцы, старые животные теряют силы, часть особей погибает. Эти изменения сопровождаются перестройкой рангов. Переходу особи в новый ранг обычно предшествуют прямые или ритуальные схватки и демонстрации, «пересмотр сил», после чего закрепляется новое положение. На роль вожака-доминанта всегда претендуют более молодые, набирающие силу и опыт животные. Они занимают его место после первого же проявления вожаком неспособности отстоять свои права.
Иерархия наиболее ярко выражена в стадах, но она характерна и для многих групп оседлых животных – семей, колоний. Иерархия может проявляться также в распределении территориальных участков у одиночных форм. Среди сусликов, охраняющих свои индивидуальные территории, доминирующие особи занимают наиболее благоприятные места в поселениях. Среди колониальных птиц (чаек, гусей, цапель и др.) наиболее крупные и опытные гнездятся в центре колонии, тогда как периферию занимают физиологически менее полноценные особи, проявляющие реакцию подчинения по отношению к доминантам.
Система доминирования-подчинения выявлена и у ряда беспозвоночных: насекомых (сверчков, жуков-чернотелок и др.), некоторых ракообразных и пр.
Установление иерархии сопровождается изменениями в гормональной системе организмов. У млекопитающих, например, возникает гипертрофия коры надпочечников. Эти изменения оказываются наибольшими у животных, находящихся в самом низу иерархической лестницы, и не проявляются у до-минантов. Стероидные гормоны, выделяемые корой надпочечников, оказывают большое влияние на процессы обмена веществ, которые, в свою очередь, лежат в основе изменения поведения.
Эффект группы. Жизнь в группе через нервную и гормональную системы отражается на протекании многих физиологических процессов в организме животного. У изолированных особей заметно меняется уровень метаболизма, быстрее тратятся резервные вещества, не проявляется целый ряд инстинктов и ухудшается общая жизнеспособность.
Оптимизация физиологических процессов, ведущая к повышению жизнеспособности при совместном существовании, получила название эффект группы. Он проявляется как психофизиологическая реакция отдельной особи на присутствие других особей своего вида. У овец вне стада учащаются пульс и дыхание, а при виде приближающегося стада эти процессы нормализуются. Одиночно зимующие летучие мыши отличаются более высоким уровнем обмена веществ, чем в колонии. Это ведет к повышенной трате энергии, истощению и часто заканчивается гибелью животных.
Затраты организмом энергии на основную жизнедеятельность учитывают, измеряя скорость потребления кислорода животными в покое. Эти показатели возрастают у всех животных, приспособленных к групповой жизни и оказавшихся вне стада, семьи или колонии. Напротив, даже простое скучивание сокращает газообмен порой до 50 %. Возможно, что это явление лежит в основе агрегации на зимовку рыб, лягушек, моллюсков, насекомых, у которых также обнаружена экономия энергетических затрат в скоплениях.
Эффект группы проявляется в ускорении темпов роста животных, повышении плодовитости, более быстром образовании условных рефлексов, повышении средней продолжительности жизни индивидуума и т. д. В группе животные часто способны поддерживать оптимальную температуру (при скучивании, в гнездах, в ульях). У многих животных вне группы не реализуется плодовитость. Голуби некоторых пород не откладывают яйца, если не видят других птиц. Достаточно поставить перед самкой зеркало, чтобы она приступила к яйцекладке. Кайры при небольшой численности пар в колонии начинают размножение только тогда, когда колония увеличится по периферии за счет других видов: чистиков, тупиков, моевок.
Эффект группы не проявляется у видов, ведущих одиночный образ жизни. Если таких животных искусственно заставить жить вместе, у них повышается раздражительность, учащаются столкновения и многие физиологические показатели сильно уклоняются от оптимума, о чем можно судить, например, по увеличению энергетических затрат на основные процессы жизнедеятельности. Так, ушастые ежи в группе повышают потребление кислорода до 134 % по сравнению с содержащимися поодиночке.
Положительный эффект группы проявляется лишь до некоторого оптимального уровня плотности популяции. Если животных становится слишком много, это грозит для всех недостатком ресурсов среды. Тогда вступают в действие другие механизмы, приводящие к снижению численности особей в группе путем ее деления, рассредоточения или падения рождаемости. Эти механизмы обсуждены ниже, при рассмотрении гомеостаза популяций.
8.5. Динамика популяций
8.5.1. Биотический потенциал
Любая популяция теоретически способна к неограниченному росту численности, если ее не лимитируют факторы внешней среды. В таком гипотетическом случае скорость роста популяции будет зависеть только от величины биотического потенциала, свойственного виду. Понятие биотического потенциала введено в экологию в 1928 г. Р. Чепменом. Этот показатель отражает теоретический максимум потомков от одной пары (или одной особи) за единицу времени, например за год или за весь жизненный цикл.
При расчетах его чаще всего выражают коэффициентом г и вычисляют как максимально возможный прирост популяции ДN за отрезок времени Дt, отнесенный к одной особи, при начальной численности популяции N0:
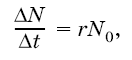
откуда
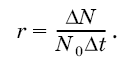
Величина биотического потенциала чрезвычайно различна у разных видов. Например, самка косули способна произвести за жизнь 10–15 козлят, трихина (Trichinella spiralis) – отложить 1,8 тыс. личинок, самка медоносной пчелы – 50 тыс. яиц, а луна-рыба – до 3 млрд икринок. Если бы все зародыши сохранялись, а все потомство выживало, численность любой популяции через определенные интервалы увеличивалась бы в геометрической прогрессии.
Кривая, отражающая на графике подобный рост популяции, быстро увеличивает крутизну и уходит в бесконечность (рис. 122). Такая кривая носит название экспоненциальной. На логарифмической шкале подобная зависимость численности популяции от времени будет представлена прямой, а биотический потенциал г отразится ее наклоном по отношению к горизонтальной оси, который тем круче, чем больше величина r.
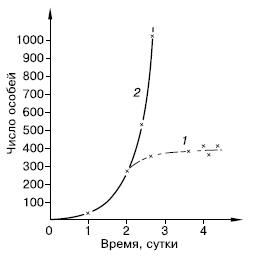
Рис. 122. Реальная (1) и теоретическая (2) кривая роста популяции парамеций
В природе биотический потенциал популяции никогда не реализуется полностью. Его величина обычно складывается как разность между рождаемостью и смертностью в популяциях: r = b – d, где b– число родившихся, а d – число погибших особей в популяции за один и тот же период времени.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 |
