

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Как видно из табл. 1, у пяти ризосферных и почвенных псевдомонад выявлена гомология с зондом czcCBAD, однако эти штаммы были чувствительны к никелю и не содержали плазмид. У восьми штаммов, выделенных из сточной воды, выявлены сигналы гибридизации с зондами czcCBAD (3 штамма), ncсСB (2 штамма) и одновременно с двумя зондами cnrXYHCBAT/ncсСB (3 штамма). Только в этой группе бактерий обнаружены 5 плазмидосодержащих штаммов.
2. Характеристика детерминанты устойчивости к кобальту/никелю в штамме Comamonas testosteroni BS501(pBS501)
2.1 Плазмидная локализация. Наибольший интерес для дальнейших исследований представлял выделенный из пробы сточной воды штамм W28, (лабораторный номер BS501(pBS501)) устойчивый к 5 мМ кобальта, 5 мМ никеля и 2 мМ цинка. В штамме обнаружена плазмида pBS501 размером около 65 т. п.н. (рис. 1, дорожка 1), которая передавалась при конъюгации в чувствительные штаммы P. putida BS394 и W340 с частотой 10–6 и обеспечивала устойчивость только к 2 мМ кобальта и никеля. Из данных Таблицы 1 видно, что при гибридизации ДНК колоний штамма BS501(pBS501) были выявлены 2 сигнала – сильный с зондом cnr и слабый с зондом ncc. Однако, гибридизация EcoRI-, HindIII - и SalI-фрагментов рестрикции ДНК pBS501 с этими зондами выявила только один положительный сигнал с опероном cnrXYHCBA, локализованным на плазмиде pMOL28.

Рис. 1. Электрофорез плазмидных ДНК из штаммов: (1) W28 (BS501(pBS501)) (65 т. п.н.), (2) W29 (90 т. п.н.); маркерные плазмиды: (3) pMG18 (102 т. п.н.), (4) pBS216 (85 т. п.н.), (5) pULB113 (68 т. п.н.).
2.2 Идентификация штамма BS501(pBS501). До настоящего времени обнаружены только три плазмиды, содержащие функциональные детерминанты устойчивости: pMOL28 (cnr) и pMOL30 (czc) в Cupriavidus metallidurans CH34 (Mergeay et al., 1985) и pTOM9 (ncc) в C. metallidurans 31a (Schmidt and Schlegel, 1994). Плазмида pBS501 (65 т. п.н.) в отличие от pMOL28 (180 т. п.н.), значительно меньшего размера и не детерминирует устойчивость к хромату и ртути. Поэтому несомненный интерес представляло выяснение таксономического положения штамма BS501(pBS501). По совокупности морфологических, физиолого-биохимических признаков и по результатам сравнительного анализа последовательности гена 16S рибосомальной РНК штамм BS501(pBS501) отнесен к роду Comamonas (класс Betaproteobacteria, порядок Burkholderiales, семейство Comamonadaceae), и идентифицирован как Comamonas testosteroni (рис. 2). В бактериях рода Comamonas описаны плазмиды, несущие гены биодеградации толуолсульфоната, сульфобензоата, хлоро - и нитробензенов, фталатов и других токсичных ксенобиотиков (Tralau et al., 2001; Wu, et al., 2006). Хотя в геномах бактерий порядка Burkholderiales выявлены последовательности, гомологичные cnr и ncc (Stoppel and Schlegel, 1995), их исследование не проводилось, и плазмиды, контролирующие устойчивость к двухвалентным катионам, в Comamonas до настоящего времени не были обнаружены.

Рис. 2. Положение штамма Comamonas testosteroni BS501(pBS501) на филогенетическом дереве порядка Burkholderiales (класс Betaproteobacteria) на основании последовательности гена 16S рРНК. Масштаб (0.01) соответствует 1 нуклеотидной замене на каждые 100 нуклеотидов. Статистическую достоверность ветвления оценивали с помощью «bootstrap-анализа» 1000 альтернативных деревьев, используя соответствующую функцию программы TREECON.
Праймеры, фланкирующие структурные гены cnrCB и cnrA оперона cnr плазмиды pMOL28, были использованы в ПЦР с плазмидной ДНК pBS501. Различия, обнаруженные в спектре и размерах ампликонов, свидетельствуют о том, что исследуемая детерминанта отличается от оперона cnr (рис. 3а, дорожки 4, 5). В то же время, положительная гибридизация с зондами cnrCB (рис. 3б, дорожка 4) и cnrA (рис. 3в, дорожка 5) указывает на их значительную гомологию.
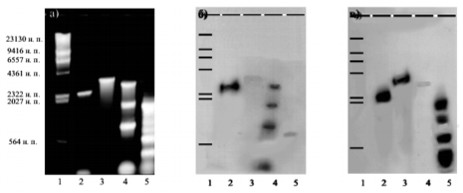
Рис. 3. Выявление гомологии детерминанты устойчивости к кобальту/никелю плазмиды pBS501 с 32Р-меченными ампликонами cnrCB (б) и cnrА (в). (а) Электрофореграмма: 1 − λHindIII; 2 и 3 − ампликоны cnrCB и cnrА, соответственно, с плазмиды cnrXYHCBA:pSUP202 («+» контроли); 4 и 5 − ампликоны «cnrCB» и «cnrА», соответственно, с плазмиды pBS501.
2.3 Индукция устойчивости к кобальту/никелю. Экспрессия оперона cnr в C. metallidurans CH34 индуцируется катионами кобальта и никеля (Schmidt and Schlegel, 1994). Предварительное выращивание инокулята штамма C. testosteroni BS501(pBS501) при 10−200 мкМ никеля или кобальта с первых часов обеспечивало быстрый рост культуры в среде, содержащей 2 мМ кобальта или никеля (рис. 4, кривые 1, 2), тогда как в вариантах без индукции наблюдалась продолжительная лаг-фаза (4−6 ч) (рис. 4, кривая 3). Из рисунка видно, что катионы цинка и кадмия не являются индукторами устойчивости (рис. 4, кривые 4 и 5).
Рис. 4. Влияние индукции системы устойчивости катионами тяжелых металлов на рост C. testosteroni BS501(pBS501) в присутствии 2 мМ никеля. Инокулят выращивали при: (1) 20 мкМ Ni, (2) 50 мкМ Со, (4) 200 мкМ Zn, (5) 50 мкМ Cd, (3) без индукции.
2.4 Устойчивость цитоплазматической мембраны клеток исходного штамма C. testosteroni BS501(pBS501) и трансконъюганта P. putida BS394(pBS501) к воздействию кобальта
С помощью метода электроориентационной спектроскопии (ЭОС) (Мирошников и др., 1986) показано, что цитоплазматическая мембрана (ЦМ) клеток C. testosteroni BS501(pBS501) и трансконъюганта P. putida BS394(pBS501), инкубированных в присутствии 50−200 мкМ кобальта, подвергается лишь незначительному повреждению. Так, при 100 мкМ кобальта для клеток C. testosteroni BS501(pBS501) и P. putida BS394(pBS501) величина Δβ/βк, характеризующая степень повреждения ЦМ, составила −0.13±0.05 и −0.15±0.05, соответственно, тогда как для чувствительного штамма P. putida BS394 Δβ/βк = −0.48±0.2. После мягкой обработки клеток штамма C. testosteroni BS501(pBS501) трипсином степень повреждения его ЦМ при воздействии 100 мкМ кобальта увеличивалась до −0.54±0.1, что может указывать на наличие защитных белков на поверхности клеток C. testosteroni BS501(pBS501). С поверхности клеток штаммов C. testosteroni BS501(pBS501) и P. putida BS394(pBS501) 5 М хлоридом лития солюбилизировались белки массой около 49, 40 и 32 кДа, которые отсутствовали у чувствительного штамма P. putida BS394 (рис. 5). Обнаруженные белки оказались близки по массе белкам, кодируемым сnr-опероном из C. metallidurans CH34: CnrC (44 кДа), CnrB (40 кДа) и CnrT (37 кДа) (Liesegang et al., 1993).
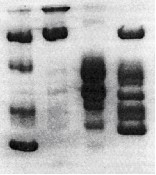
Рис 5. Электрофореграмма белков, солюбилизированных 5 М хлоридом лития с поверхности клеток устойчивых штаммов, в 9% ПААГ-ДСН: (1) белковые маркеры (кДа), (2) P. putida BS394, (3) C. testosteroni BS501(pBS501), (4) P. putida BS394(pBS501).
Измерения ЭОС клеток проводились при 20°С через 15 мин после внесения в бактериальную суспензию кобальта. Поэтому обнаруженные поверхностные белки могли участвовать в процессах экспорта катионов тяжелых металлов из клеток исследуемых устойчивых бактерий, однако не исключено также их участие в биосорбции и иммобилизации металлов.
3. Характеристика штаммов PGPR Pseudomonas, содержащих плазмиду pBS501
3.1 Получение устойчивых вариантов PGPR. Плазмида pBS501 с помощью конъюгации была перенесена в ризосферные штаммы P. aureofaciens BS1393, P. fluorescens 38a и P. chlororaphis PCL1391. С частотой 10−4−10−5 получены трансконъюганты, в которых плазмида стабильно сохраняется в неселективных условиях, как минимум, в течение 10 пассажей и обеспечивает повышение устойчивости к кобальту (0.5−2.0 мМ) от 5 до 20 раз, и никелю (1.0−2.5 мМ) от 2 до 10 раз (табл. 2).
Таблица 2. Уровни устойчивости к тяжелым металлам штаммов Pseudomonas и Comamonas в трис-минеральной среде с глютаматом натрия или глюкозой
Штаммы | Максимальный уровень устойчивости (мМ) | |||
Co | Ni | Zn | Cd | |
P. aureofaciens BS1393 | 0.1 | 1.0 | 1.5 | 0.2 |
P. aureofaciens BS1393(pBS501) | 2.0 | 2.0 | 1.5 | 0.2 |
P. chlororaphis PCL1391 | 0.05 | 0.15 | 2.5 | 0.2 |
P. chlororaphis PCL1391(pBS501) | 0.5 | 2.0 | 2.5 | 0.2 |
P. fluorescens 38a | 0.05 | 0.1 | 2.0 | 0.1 |
P. fluorescens 38a(pBS501) | 0.5 | 1.0 | 2.0 | 0.1 |
C. testosteroni B-1241 | 0.5 | 1.0 | 1.0 | 0.1 |
C. testosteroni B-1241(pBS501) | 5.0 | 5.0 | 1.0 | 0.1 |
C. acidovorans B-1251 | 1.0 | 2.0 | 1.0 | 0.2 |
C. acidovorans B-1251(pBS501) | 5.0 | 5.0 | 1.5 | 0.2 |
C. testosteroni BS501(pBS501) | 5.0 | 5.0 | 1.0 | 0.1 |
3.2 Супрессия фитопатогенов и продукция индольных соединений ризосферными штаммами в присутствии кобальта/никеля. Исходные ризосферные штаммы являются продуцентами фитогормона индолил-3-уксусной кислоты (ИУК) и противогрибных антибиотиков – феназинов (P. aureofaciens BS1393), пиолютеорина (P. fluorescens 38a) и феназинкарбоксамида (P. chlororaphis PCL1391). Известно о положительном влиянии микромолярных концентраций тяжелых металлов на синтез ИУК (Belimov et al., 2000, Белимов, 2008) и некоторых антибиотиков (Slininger and Jackson, 1992; Chin-A-Woeng et al., 1998 Duffy and Defago, 1999). В настоящей работе показано, что в культуральной среде устойчивых штаммов P. aureofaciens BS1393(pBS501) и P. chlororaphis PCL1391(pBS501), выращенных в присутствии 2 мМ никеля или кобальта, концентрация индольных соединений (1.6−2.0 мкг/мг биомассы) в 1.5−2 раза выше по сравнению с чувствительными вариантами.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
