

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ЦИКЛ УГЛЕРОДА
Цикл углерода в почве — важный геохимический цикл, обеспечивающий стабильность углерод-кислородной системы Земли. С круговоротом углерода связаны важнейшие процессы — фотосинтез и судьба почвенного органического вещества. От динамики этих процессов зависит состояние наземного покрова и атмосферы.
В круговороте углерода и кислорода наиболее четкое выражение находит взаимозависимость и связь всех живых существ на Земле. Благодаря участию в этом цикле микроорганизмов почвы как деструктивного звена происходит замыкание круговорота и совершается циклическое превращение веществ и энергии в биосфере. В этом главнейшая функция почвенных микроорганизмов.
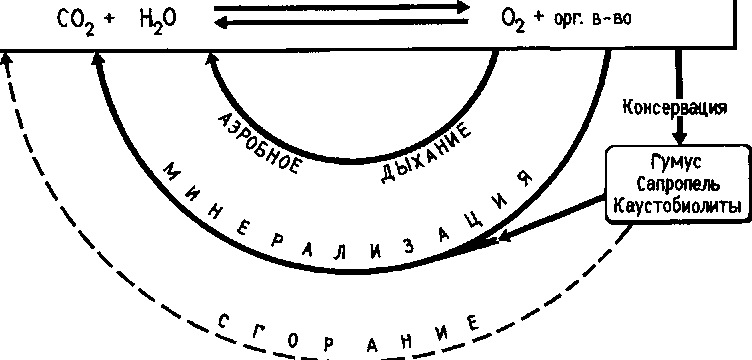
Рис. 68. Цикл углерода и кислорода
В цикле углерода можно выделить два звена, имеющих планетарные масштабы и связанных с выделением и поглощением кислорода:
1) связывание С02 в процессе фотосинтеза и генерация кислорода;
2) минерализация органических веществ с выделением С02 и затратой кислорода.
Первый процесс совершается в основном растениями, второй — микроорганизмами (рис. 68).
Углерод в составе биосферы представлен разнообразными формами — от простейших одноуглеродных соединений до сложнейших молекул лигнина. Рассмотрим разные механизмы синтеза углеродных соединений с участием микроорганизмов почвы и разложения органических веществ в ряду от наиболее простых к сложным и стабильным формам углеродных соединений.
Процессы связывания (фиксации) С02
Из углекислоты создается вся первичная органическая продукция. Содержание С02в атмосфере около 0,03%, в почвенном воздухе — на порядок больше. Биологическое связывание С02 происходит в процессах фотосинтеза, хемосинтеза и гетеротрофной фиксации. Фотосинтез в основном происходит в наземном слое биосферы, два последних процесса — преимущественно в почве.
Фотосинтез. Фиксация С02 в фотосинтезе происходит за счет световой энергии:
С02 + Н20 + hv -> (СН20) + 02,
где hv — световая энергия.
На этот процесс в биосфере расходуется 0,1% энергии Солнца, попадающей на Землю. Запасенная в продуктах фотосинтеза энергия служит важным энергетическим источником для ряда процессов, происходящих на Земле. В результате фотосинтеза ежегодно создается 180 млрд т органического вещества, из них 120 млрдт на суше и 60 млрд т в океане и выделяется приблизительно 100 млрд т свободного 02. Фотосинтез осуществляется высшими растениями, водорослями и некоторыми бактериями. В фотосинтезе участвуют пигменты, содержащиеся в хлоро - пластах (у растений и водорослей) или в мембранных структурах бактерий — хлоросомах, тилакоидах, фикобилисомах.
В основе фотосинтеза лежат окислительно-восстановительные процессы, в которых электроны переносятся от донора-восстановителя к акцептору. Восстановителем в растительном фотосинтезе выступает молекула воды, акцептором — С02. При этом образуется органическое вещество и выделяется кислород. У бактерий донорами электрона могут быть сероводород, сера, водород, органические соединения. Кислород в этом процессе не выделяется (бескислородный фотосинтез).
Фиксация С02 у автотрофных организмов происходит за счет универсального механизма, известного как цикл Кальвина, или рибулозобифосфатный цикл. У зеленых бактерий имеется циклический механизм фиксации С02, отличный от цикла Кальвина. В его основе лежат реакции восстановительного карбоксилирова - ния органических кислот, получившие название цикла Арнона.
Фототрофные прокариоты составляют группу микроорганизмов с разным отношением к кислороду — от строгих анаэробов до аэробов, у которых кислород образуется внутриклеточно. Есть факультативные анаэробы, аэротолерантные формы и микро - аэрофилы.
Из фототрофных прокариот в почвах обитают цианобакте - рии, галобактерии (на солончаках) и пурпурные несерные бактерии в болотах и на рисовых полях. Некоторые фототрофные прокариоты развиваются на мхах, папоротниках, высших растениях, они также вступают в более тесные ассоциации с другими организмами, например цианобактерии с грибами образуют некоторые лишайники.
Хемосинтез. Фиксация С02 в хемосинтезе происходит за счет энергии окисления внешнего неорганического донора электрона. Этот процесс впервые был описан в конце прошлого века и назван аноргоксидацией. Хемоавтотрофы известны только среди бактерий (прокариот). К хемосинтезиру - ющим организмам относятся нитрификаторы, серобактерии, железобактерии, водородные бактерии, карбоксидобактерии. Они называются так по субстратам окисления, которыми могут быть NH3, NO2", H2S, S, Fe2+, H2, CO. Некоторые виды — облигатные хемолитоавтотрофы, другие — факультативные. К последним относятся карбоксидобактерии и водородные бактерии. В отсутствие органических веществ они используют энергию окисления водорода или окиси углерода для фиксации С02, а при наличии органических соединений переходят на гетеротрофный тип питания, т. е. они являются миксотрофами.
Гетеротрофная фиксация С02. Помимо фото - и хемосинтеза, где углекислота служит единственным источником углерода, она может ассимилироваться гетеротрофно, путем присоединения к готовым углеродным цепям. К гетеротрофному связыванию углекислоты способны и такие фотоавтотрофы как высшие растения, а также животные. Акцепторами С02 могут быть различные органические кислоты, чаще всего, пировиноградная (реакция Вуда-Веркмана):
СНзОСООН + С02 СООНСН2СОСООН,
хотя всего известно до 10-12 способов включения С02 при гетеротрофной фиксации.
До 3-10% углерода биомассы гетеротрофных организмов может быть построено из С02, как это было показано в опытах при использовании меченой |4С02.
Другие пути превращения одноуглеродных соединений.
Образование метана биологическим путем из С02 и других одноуглеродных источников — метановое брожение — происходит в болотах, торфяниках, иловых отложениях озер, метантен - ках, рубце жвачных животных, кишечном тракте человека. В почве этот процесс протекает там, где возникают анаэробные условия, и образуется водород в первичных процессах анаэробного превращения органических веществ.
Метанобразующие бактерии (метаногены) — вторичные анаэробы, так как они перерабатывают продукты, поступающие от других анаэробов. Метан образуется за счет восстановления С02 водородом по следующей схеме:
С02 + 4Н2 = СН4 + 2Н20.
Восстановлению могут подвергаться также СО, муравьиная кислота и метанол.
В газовой атмосфере торфяников метан часто превышает 30%. Группа метанобразующих бактерий гетерогенна. Среди них есть кокковидные, палочковидные, сарциноподобные организмы и спириллы. Все метаногенные бактерии относятся к архебакте - риям, их родовые названия начинаются со слова methan (Methano - coccus, Methanosarcina, Methanobacterium, Methanospirillum). Метанобразующие бактерии строгие анаэробы. Кислород убивает их, с чем сопряжены большие трудности при работе с ними в лаборатории. Необходимо применять специальные методы, которые были разработаны в последние 20 лет, и было проведено подробное их изучение. В ряде анаэробных экосистем основным субстратом для образования метана служит ацетат. Метанобразующие бактерии составляют последнее звено анаэробной пищевой цепи, в начале которой находятся полисахариды (целлюлоза, крахмал, хитин), липиды, белки. В этой цепи участвуют: 1) бактерии, сбраживающие целлюлозу до сукцината, пропионата, бутирата, лактата, ацетата, спиртов, С02 и Н2; 2) ацетогенные бактерии, сбраживающие эти первичные продукты до ацетата, формиата, С02 и Н2.
Метанобразующие бактерии находятся в симбиотических (мутуалистических) взаимоотношениях с бактериями, образующими газообразный водород. Первые не могут развиваться без водорода, а вторые сами себя отравляют избытком образуемого ими водорода. При совместном развитии этих двух групп бактерий свободный водород практически не обнаруживается, так как он моментально используется метанотрофами.
Метанобразующие бактерии активируют водород и осуществляют его окисление, сопряженное с восстановлением С02. Клеточное вещество они также строят из С02, т. е. являются хемоав - тотрофами. Они осуществляют карбонатное дыхание (в качестве источника С02 используют СаС03). Ферменты метанобразующих бактерий отличаются большой специфичностью. Образование метана может происходить из ацетата или С02
Метан, помимо биологического пути, образуется и в результате геохимических процессов. Он составляет основную часть природных углеводородных газов.
Окисление метана (метанотрофы) — биологический процесс, активно протекающий в районах метаногенеза — в местах газовых и нефтяных месторождений, в переувлажненных почвах, в поверхностной пленке болот. Метанокисляющие бактерии перехватывают метан, который образуется в анаэробной зоне, и окисляют его до С02 через метанол, формальдегид и муравьиную кислоту:
СН4 СН3ОН -> СНОН -> НСООН С02.
Этот путь диссимиляции (катаболизма) осуществляется с участием ферментов цепи переноса электронов. Ассимиляция углерода метана происходит на уровне формальдегида несколькими путями, отличными от автотрофной фиксации С02 в цикле Кальвина. Микроорганизмы, окисляющие метан, получили название метанотрофов. Среди них есть облигатные формы, использующие в качестве углеродного источника питания метан или его производные (метанол и метиламины) и факультативные мети - лотрофы, ассимилирующие как одноуглеродные соединения (метанол, формальдегид, метиламин, но не метан), так и другие органические вещества. Облигатные метанотрофы — группа ме - танокисляющих бактерий семейства Methylomonadaceae, которые относят к родам Methylomonas, Methylobacillus, Methylococcus, Methylosinus и др. Хотя формально их следует считать гетеротро - фами (метан — органическое вещество), но по многим морфологическим и физиологическим признакам они близки к таким литоавтотрофам, как нитрифицирующие бактерии семейства Nitrobacteriaceae. Процесс окисления метана сопровождается у них окислением аммиака до нитритов, они имеют хорошо развитую систему внутриклеточных цитоплазматических мембран, пути метаболизма метанокисляющих бактерий сходны с таковыми ав - тотрофных нитратных бактерий.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
