

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
БАВ можно разделить на четыре группы в зависимости от способности вызывать биологический ответ: агонисты, частичные агонисты, антагонисты и неактивные соединения. К агонистам относятся соединения, способные вызывать максимальный ответ, однако необходымые для этого концентрации могут быть различны.
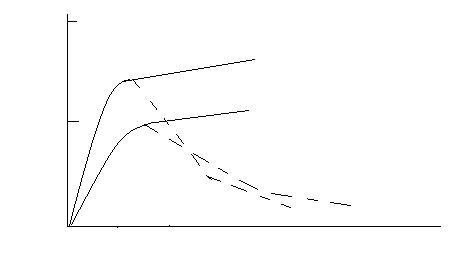
Рис. БАВ можно разделить на следующие группы: агонисты (А), частичные агонисты (Б), антагонисты (А+В и Б+В) и неактивные агенты (Г).
По оси абсцисс – обратный логарифм концентрации вещества. По оси ординат - % от максимального ответа.
Частичные агонисты вызывают ослабленный ответ, даже если используются в очень высокой концентрации. Антагонисты обычно не оказывают эффекта сами по себе, но послностью ингибируют действие агонистов и частичных агонистов. Большая группа соединений, структурно сходных с активными БАВ, не обладает собственным эффектом и не влияет на действие агонистов и антагонистов. Их относят к категории неактивных веществ.
Частичные агонисты нередко конкурируют с агонистами за связывание с рецептором (и активацию его), в этих случаях они становятся частичными антагонистами. Степень торможения активности агониста, вызываемая частичными или полными антагонистами, зависит от соотношения концентраций соответствующих БАВ. Как правило, антагонист вызывает торможение в концентрации намного большей, чем та, в которой агонист оказывает максимальный эффект.
Изучение рецепторов (от лат. слова receptio - восприятие) стало в последнее время одним из важнейших направлений клеточной биологии. Рецепторы играют исключительно важную роль жизнедеятельности клетки, в понимании механизмов гомеостаза и патогенеза различных заболеваний, для понимания механизмов действия эндогенных и экзогенных БАВ, лекарственных средств и для направленного поиска новых лекарственных соединений
Рецептор - это любая биомолекула, специфически связывающая химическое соединение (лиганд, лекарственное вещество, агонист, антагонист) на поверхности или внутри клетки и трансформирующая полученную информацию в биологический ответ. Биологический эффект БАВ (лигандов) начинается с их связывания со специфическими рецепторами, а завершается, как правило, диссоциацией лиганда и рецептора.
Рецепторы представляют собой материальные субстраты чувствительности и реактивности клеток. Рецепторами могут быть различные клеточные структуры, которые ответственны за реакцию клетки на гормональный сигнал и проведение его внутрь клетки. Они представляют собой первичную ступень активации каскада биохимических реакций, конечным итогом которых является определённый данным биорегулятором функциональный ответ клетки. Основой специфичности связывания лекарство-рецептор является их взаимная комплементарность.
Клетку-мишень определяют по способности избирательно связывать данный лиганд с помощью такого рецептора, причем для количественной оценки взаимодействия используют радиоактивные лиганды.
Количество рецепторов в клетке или на ее поверхности находится в динамическом состоянии: оно регулируется физиологически и изменяется при заболеваниях или под влиянием терапевтических средств. Концентрация рецепторов на мембране и сродство к лиганду (аффинность) являются регулируемыми параметрами. Изменение этих параметров происходит очень быстро и существенным образом сказывается на чувствительности клетки к лиганду. Например, в клетках, подвергнутых воздействию ?-адренергических агонистов, в течение некоторого времени (от нескольких минут до часов) в ответ на новое добавление агониста прекращается активация аденилатциклазы и исчезает биологический ответ. Такая десенситизация опосредуется двумя механизмами. Первый включает утрату рецепторов плазматической мембраной. Эта понижающая регуляция осуществляется путем секвенирования (связывания) рецепторов в клетке, т. е. отделения их других компонентов системы клеточного ответа. В частности от регуляторной и каталитической субъединиц аденилатциклазы. После удаления агониста рецепторы возвращаются на поверхность клетки и чувствительность к лиганду восстанавливается. Второй механизм десеситизации ?-адренергической системы – ковалентная модификация рецепторов путем фосфорилирования. Это сАМФ-зависимый процесс, который не сопряжен с изменением числа рецепторов и их перемещением. Некоторые лиганды (ангиотензин II и пролактин) осуществляют повышающую регуляцию своих рецепторов. Эти изменения количества рецепторов могут происходить очень быстро (за время, измеряемое минутами или часами) и служат важным средством регуляции биологического ответа.
В процессе рецепции важная роль принадлежит мембранным липидам, образующим более или менее прочные связи с рецепторными белками. Связывание с липидами включает каскад внутриклеточных сопряжённых реакций. Липиды не только принимают участие в узнавании рецепторами лигандов, оказывая регулирующее влияние на конформацию рецепторов, но и сами выступают в роли рецепторов.
Клеточные компоненты, вовлеченные в процесс начального действия лекарств, обычно определяют как рецепторную субстанцию или просто рецептор. Химические группы, которые участвуют в комбинации вещество-рецептор, и прилегающие к ним структуры, облегчающие связывание лекарства с рецептором, известны как рецепторные области. Важно отметить, что кроме связывания вещества с рецептором, вызывающим эффект, существуют и другие типы связывания. Например, лекарство может связываться с альбумином - белком плазмы крови, который переносит лекарство по всему организму, - или с ферментами, участвующими в биотрансформации веществ в тканях. В результате связывания не происходит фармакологического эффекта, поэтому такие рецепторы получили название "молчащих" или вторичных рецепторов.
Взаимодействие рецептора и лекарства определяется законом действующих масс, что подчеркивает кинетический характер взаимодействия. Из классической теории рецепторов следует, что этот эффект пропорционален фракции рецепторов, связанных с лекарственным веществом, и максимальный эффект развивается тогда, когда все клеточные рецепторы заняты лекарством. Впоследствии в эти представления была внесена поправка. Было показано, что максимальный эффект развивается лишь после того, как критическая часть рецепторов занята лекарством. В качестве рецепторов могут выступать карбоксильные группы белка, аминогруппы, сульфгидрильные группы, остатки фосфорной кислоты. С рецепторными группами лекарство может связываться при помощи ионной, водородной, ван-дер-ваальсовых связей, а также ковалентных связей, последний вид связей наиболее прочный. При большинстве взаимодействий имеется комбинация целого ряда связей.
Наличие у рецепторов физиологически активных веществ специфичности было постулировано ещё Эрлихом. Отражением специфичности рецепторов является их гетерогенность, выявляемая фармакологически. Биологическая специфичность межмолекулярных взаимодействий вначале была продемонстрирована не для лиганд-рецепторных комплексов, а для фермент-субстратных взаимодействий, когда с помощью ренгеноструктурного анализа была показана высокая комплементарность активного центра ряда ферментов и их субстратам и ингибиторам. Для биологической специфичности можно выделить два компонента: термодинамический (связывание за счёт простых молекулярных взаимодействий) и кинетический (проявление некоторой реакции как результат взаимодействия молекул-партнёров). Рассмотрим более подробно те методические подходы, которые применяются для выявления специфичности рецепторов.
После успехов в идентификации рецепторов БАВ и развития радиолигандной техники стало ясно, что все положения о специфичности ферментов относятся также и к рецепторам БАВ. Чаще всего изучается связывание радиолигандов с мембранными структурами, содержащими рецепторы. В микроплейтах инкубируют мембраны с радиолигандами. Для отделения мембран, связавших радиолиганд, содержимое лунок фильтруют и затем определяют величину радиоактивности мембран (при этом величина связанного радиолиганда отражает общее число связывающих участков как специфических, так и неспецифических. В другой серии лунок радиолиганд и мембраны инкубируют в присутствии большого избытка немеченого лиганда. Исходя из того, что количество специфических участков намного меньше неспецифических в присутствии избытка немеченого лиганда, они полностью оккупируются им. Таким образом, различие между специфическим связыванием радиолиганда и общим связыванием представляет собой величину насыщаемого, специфического или вытесняемого конкурентом связывания лиганда с мембранами. Неспецифическое связывание радиолиганда можно уменьшить путём промывания лиганд-рецепторного комплекса холодным буфером.
Математическое описание взаимодействия рецептор-лиганд
Целесообразно рассмотреть общие вопросы рецептологии, которые являются базой изучения механизма действия лекарственных веществ (ЛВ). Первые попытки описать математически взаимодействие ЛВ с рецепторами, основываясь на данных фармакологических экспериментов (влияние ацетилхолина на мышечные клетки), предпринял Кларк, предложивший простую оккупационную теорию. Однако оказалось, что некоторые вещества (антагонисты), хотя и связываются с рецепторами, но не вызывают эффекта. Учитывая этот факт Ариенс усовершенствовал концепцию, предложенную Кларком, и создал сложную оккупационную теорию. Обобщённые её положения носят название «классической теории действия ЛВ».
Классическая теория основана на 2-х постулатах:
Величина биологического эффекта прямо пропорциональна концентрации комплексов ЛВ-рецептор [ЛР]:Е = ?[ЛР]
Максимальный эффект имеет место при оккупации всех рецепторов [R].Классическая теория действия ЛВ основывается на законе действия масс и описывается уравнением аналогичным описанию кинетики взаимодействия субстрата с ферментом, если продукт не образуется
Еmax [Ло]
Е = , где
[Ло] + Kд
Е - биологический эффект, Kд - константа диссоциации комплекса ЛР, [Ло] - общая концентрация вещества, Еmax - максимальный биологический эффект вещества.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 |
