

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Лимитирующий фактор — любое условие, приближающееся к пределу толерантности или превышающее его. Лимитирующим фактором может быть не только недостаток, но и избыток таких, например, факторов, как тепло, свет и вода.
Экосистема — основная функциональная единица в экологии. Её компоненты (и организмы, и неживая среда) взаимно влияют на свойства друг друга и необходимы для поддержания жизни в той её форме, которая существует на Земле.
Антропогенные экологические факторы — факторы, связанные с влиянием человека на жизнедеятельность экосистемы.
ЛЕКЦИЯ №5 ПРОСТЕЙШИЕ МАТЕМАТИЧЕСКИЕ МОДЕЛИ ПОПУЛЯЦИОННОЙ ДИНАМИКИ. ИССЛЕДОВАНИЕ ОДНОРОДНОЙ ПОПУЛЯЦИИ, ЖИВУЩЕЙ ИЗОЛИРОВАННО В НЕИЗМЕННОЙ СРЕДЕ
ПРОСТЕЙШИЕ МАТЕМАТИЧЕСКИЕ МОДЕЛИ ПОПУЛЯЦИОННОЙ ДИНАМИКИ
При анализе любой экологической системы главная проблема — выбор существенных переменных. Рассматривая математические модели роста популяций, сконцентрируем внимание на численности или плотности особей популяции и на этой основе разработаем соответствующую динамическую теорию. Впервые такой подход использовал В. Вольтерра, с тех пор им пользуются большинство экологов.
Плотность популяции — это величина популяции, отнесенная к единице пространства. Её измеряют и выражают числом особей или биомассой популяции на единицу площади или объема, например, 700 деревьев на 1 га или 300 кг рыбы на 1 га поверхности водоема.
Исследуем простейшие модели, описывающие изменения во времени только общей численности популяции N. При построении этих моделей все особи, составляющие популяцию, совершенно идентичны, а величина численности принимается исчерпывающей характеристикой популяции как динамического объекта. Даже при таком сравнительно простом описании популяции можно построить модели, отражающие ряд экспериментальных фактов, касающихся роста популяций.
Некоторые из рассмотренных моделей в дальнейшем будем использовать для решения прикладных задач, связанных с численностью эксплуатируемых популяций.
ИССЛЕДОВАНИЕ ОДНОРОДНОЙ ПОПУЛЯЦИИ, ЖИВУЩЕЙ ИЗОЛИРОВАННО В НЕИЗМЕННОЙ СРЕДЕ
Биологические сообщества состоят из нескольких популяций биологических видов, живущих в общей среде. Обычно индивидуумы этих сообществ оспаривают одну и ту же пищу или одни виды живут за счет других, которыми они питаются. Они могут оказывать друг другу помощь. Всё это входит в общее явление борьбы за существование. Количественный характер этого явления проявляется в заданной среде в виде изменений численности индивидуумов, составляющих разные популяции. При одних условиях эти изменения состоят из флуктуации вокруг средних значений, при других — сводятся к исчезновению или прогрессирующему увеличению некоторых видов.
Для того чтобы охарактеризовать одним единственным числом некоторую популяцию в конкретной области, предположим, что все особи в популяции одинаковы и тип индивидуума не меняется со временем. Будем считать, что исследуемая популяция сосуществует с другими видами без прямого или косвенного взаимного влияния в неизменной среде, представляющей всегда одни и те же возможности максимально благоприятного существования для этой изолированной популяции.
Если вместо разрывных целочисленных функций, описывающих численность особей, введем непрерывные дифференцируемые функции, имеющие в каждый момент времени ту же целую часть, что и разрывные, то для короткого интервала времени заданной длины в достаточно многочисленной популяции число рождений и число смертей пропорциональны общей численности индивидуумов, существующих в данный момент. Если предположить, что численность популяции изменяется непрерывно, так что поколения пересекаются, а скорость прироста dN индивидуумов в малом интервале времени пропорциональна численности N популяции, то, приписывая это свойство функции N = N(t), рассматриваемой как непрерывная, получаем
![]()
где е — коэффициент, называемый относительной или удельной скоростью роста популяции, представляющий собой разность между коэффициентом рождаемости В (т. е. скоростью рождаемости в единицу времени на одну особь) и коэффициентом смертности D:
![]()
Простейшая модель (2.1) была предложена Мальтусом (для роста населения Земли) . Если ![]() , то модель ведёт к экспоненциальному (т. е. очень быстрому) росту N с течением времени:
, то модель ведёт к экспоненциальному (т. е. очень быстрому) росту N с течением времени:


![]()
где N0 — численность популяции в начальный момент. Это известный закон Мальтуса — экспоненциальный рост численности популяции в неограниченной среде. Такая динамика численности характерна для начальной фазы роста колонии бактерий, когда все необходимые для роста питательные вещества находятся в избытке.
Утверждение, что в неограниченной стационарной и благоприятной среде размер популяции экспоненциально возрастает — одно из основных экологических принципов.
Однако экспоненциальный закон роста никогда не проявляется в полной мере благодаря воздействию окружающей среды (в благоприятных условиях потомство одной пары мух через несколько лет весило бы больше, чем земной шар).
Коэффициент относительной скорости роста е, называемый также коэффициентом прироста популяции, характеризует внутренне присущую живым организмам способность к увеличению численности в отсутствие лимитирующих факторов среды. Показатель е используется также для количественного выражения «репродуктивной приспособленности» в генетическом смысле. Величину е часто называют мальтузианским параметром популяции.
Показатель роста е можно вычислить экспериментально по двум измерениям численности популяции на фазе нелимитируемого роста этой популяции. Если N1 и N2 — число индивидуумов популяции в моменты t1 и t2 , то из (2.3) следует
![]()
При ![]() уравнение (2.1) справедливо лишь для ограниченного периода времени; в конечном счёте растущая популяция исчерпает наличные ресурсы. В природных условиях, где ресурсы, обеспечивающие рост, всегда ограничены, эффект безграничного экспоненциального роста не наблюдается. Всегда существует предельная численность К, которой может достигнуть популяция в условиях ограниченности ресурса (величину К обычно называют «ёмкостью» среды):
уравнение (2.1) справедливо лишь для ограниченного периода времени; в конечном счёте растущая популяция исчерпает наличные ресурсы. В природных условиях, где ресурсы, обеспечивающие рост, всегда ограничены, эффект безграничного экспоненциального роста не наблюдается. Всегда существует предельная численность К, которой может достигнуть популяция в условиях ограниченности ресурса (величину К обычно называют «ёмкостью» среды): ![]() . Когда численность популяции становится слишком большой, мальтусовская жёсткая модель с постоянным коэффициентом е перестаёт быть применимой. При слишком больших N конкуренция за ресурсы (пищу, пространство и т. п.) приводит к уменьшению е, и жёсткая модель Мальтуса должна быть заменена мягкой моделью с зависящим от N коэффициентом прироста популяции
. Когда численность популяции становится слишком большой, мальтусовская жёсткая модель с постоянным коэффициентом е перестаёт быть применимой. При слишком больших N конкуренция за ресурсы (пищу, пространство и т. п.) приводит к уменьшению е, и жёсткая модель Мальтуса должна быть заменена мягкой моделью с зависящим от N коэффициентом прироста популяции 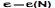 .
.
Первая модель, учитывающая этот факт, была предложена в 1825 г. Б. Гомпертцем:
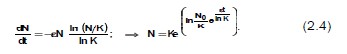
![]()
Заметим, что эта модель описывает эффект «насыщения», но эксперименты с животными показали, что этот эффект наступает гораздо быстрее, чем следует из модели Гомпертца.
ЛЕКЦИЯ № 6 ЛИМИТИРОВАННАЯ ПОПУЛЯЦИЯ С ЛОГИСТИЧЕСКИМ ЗАКОНОМ РОСТА. ОБОБЩЕННАЯ ЛОГИСТИЧЕСКАЯ ПОПУЛЯЦИЯ
ЛИМИТИРОВАННАЯ ПОПУЛЯЦИЯ С ЛОГИСТИЧЕСКИМ ЗАКОНОМ РОСТА. ОБОБЩЕННАЯ ЛОГИСТИЧЕСКАЯ ПОПУЛЯЦИЯ
В настоящее время существует много популяционных моделей с разными законами локального роста. Например, если предположить, что коэффициенты рождаемости В и смертности D в (2.2) могут зависеть от N, считая, что от пространственных координат они не зависят, то уравнение (2.1) в силу (2.2) записывают следующим образом:

Для многих видов функция рождаемости В определяется лишь физиологическими пределами рождаемости и не зависит от N, так что В(N) = n = const, где n — естественная рождаемость (или плодовитость).
Что касается зависимости функции смертности D от N, то практически для всех популяций D(N) — монотонно возрастающая функция, причем D(0) = m > 0 — естественная смертность, а возрастание смертности с ростом N объясняется ростом конкуренции за определённый ресурс (питание, пространство и т. п.).
Рассмотрим подробно этот тип популяции. Наиболее простая форма зависимости D(N) — линейная: ![]() . Тогда
. Тогда
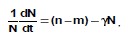
Откуда
![]()
где ![]() — естественная скорость роста в отсутствие
— естественная скорость роста в отсутствие
лимитирования, ? — коэффициент лимитирования,
соответствующий потребности популяции в пище и называемый иногда «коэффициентом прожорливости»; слагаемое ![]() в правой части (2.6) называют фактором тесноты. Это уравнение записывают в виде
в правой части (2.6) называют фактором тесноты. Это уравнение записывают в виде

и называют логистическим уравнением или логистической моделью. Интегрируя логистическое уравнение, получаем
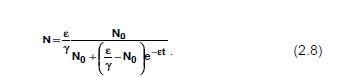
Популяция с таким законом роста называется логистической. Отметим некоторые свойства решения (2.8). Во-первых, из (2.8)
следует, что

т. е. численность популяции стремится к постоянной величине, которая прямо пропорциональна удельной скорости роста популяции в нелимитированной среде е и обратно пропорциональна коэффициенту лимитирования ? При этом возможны два случая: ![]() ?. Различия между ними показаны на рис. 2.1.?Отметим, что соотношение (2.8) описывает, в частности, популяции фруктовых вредителей и некоторых видов бактерий. Рост биомассы дрожжевых клеток в культуре и кривая роста, предсказанная логистическим уравнением, изображены на рис. 2.2.
?. Различия между ними показаны на рис. 2.1.?Отметим, что соотношение (2.8) описывает, в частности, популяции фруктовых вредителей и некоторых видов бактерий. Рост биомассы дрожжевых клеток в культуре и кривая роста, предсказанная логистическим уравнением, изображены на рис. 2.2.
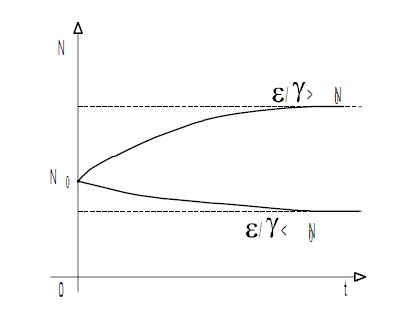
Рисунок 2.1 — Кривые изменения численности логистической популяции
Отношение (2.9) характеризует чувствительность вида к нехватке пищи: если дробь ![]() мала, то вид очень чувствителен к нехватке пищи. Во-вторых, из (2.8) следует, что при малых N0 и конечном t
мала, то вид очень чувствителен к нехватке пищи. Во-вторых, из (2.8) следует, что при малых N0 и конечном t
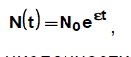
т. е. в популяции малой численности (когда отсутствует заметное влияние лимитирующих факторов) кривая изменения численности
сходна с кривой динамики роста (2.3) в отсутствие лимитирующих
факторов среды.
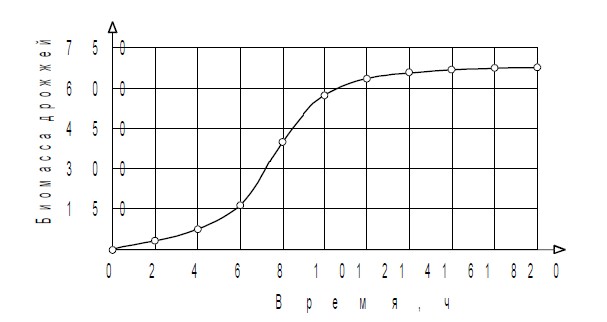
Рисунок 2.2 — Кривая роста биомассы дрожжевых клеток

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
