

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
- Повышение концентрации Са2+ в цитозоле клетки увеличивает скорость взаимодействия Са2+ с неактивным цитозольным ферментом протеинкиназой С (ПКС) и белком кальмо-дулином, таким образом сигнал, принятый рецептором клетки, раздваивается. Связывание протеинкиназы С с ионами кальция позволяет ферменту вступать в кальций-опосредованное взаимодействие с молекулами "кислого" фосфолипида мембраны, фосфатидилсерина (ФС). Диацилглицерол, занимая специфические центры в протеинкиназе С, ещё более увеличивает её сродство к ионам кальция. На внутренней стороне мембраны образуется ферментативный комплекс - [ПКС][Са2+] [ДАГ][ФС] - активная протеинкиназа С, фосфорилирующая специфические ферменты по серину и треонину.
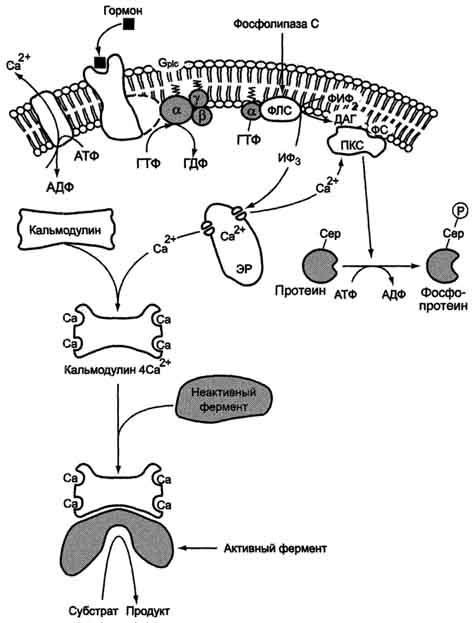
Рис. 5-42. Инозитолфосфатная система.
Участие белка кальмодулина в инозитолфосфатной передаче сигнала
В клетках многих тканей присутствует белок кальмодулин, который функционирует как внутриклеточный рецептор Са2+, он имеет 4 центра для связывания Са2+. Комплекс [кальмодулин] -[4 Са2+] не обладает ферментативной активностью, но взаимодействие комплекса с различными белками и ферментами приводит к их активации.
Как и большинство систем трансмембранной передачи сигналов, инозитолфосфатная система имеет не только механизм усиления, но и механизм подавления сигнала. Присутствующие в цитозоле инозитол-1,4,5-трифосфат (ИФ3) и диацилглицерол (ДАТ) в мембране могут в результате серии реакций опять превращаться в фосфатидилинозитол-4,5-бисфосфат (ФИФ2). Ферменты, катализирующие восстановление фосфолипида, активируются фосфорилированием протеинкиназой С.
Концентрация Са2+ в клетке снижается до исходного уровня при действии Са2+-АТФ-аз цито-плазматической мембраны и ЭР, а также Na+/Ca2+-и Н+/Са2+-транслоказ (активный антипорт) клеточной и митохондриальной мембран.
- Функционирование транслоказ Са2+ и Са2+-АТФ-аз может активироваться: комплексом [кальмодулин] [4 Са2+]; протеинкиназой А (фосфорилированием); протеинкиназой G (фосфорилированием). Понижение концентрации Са2+ в клетке и диацилглицерола в мембране приводит к изменению конформации протеинкиназы С, снижению её сродства к фосфатидилсерину, фермент диссоциирует в цитозоль (неактивная форма). Фосфорилированные протеинкиназой С ферменты и белки под действием фосфопротеинфосфатазы переходят в дефосфорилированную форму.
Передача сигнала с помощью внутриклеточных рецепторов
Передача сигнала липидорастворимых стероидных гормонов и тироксина возможна только при прохождении этих гормонов через плазматическую мембрану клеток-мишеней (рис. 5-43).
Рецепторы гормонов могут находиться в цитозоле или в ядре. Цитозольные рецепторы связаны с белком-шапероном (часто это группа белков-шаперонов). Ядерные и цитозольные рецепторы стероидных и тиреоидных гормонов содержат ДНК-связывающий домен, характеризующийся наличием двух структур "цинковых пальцев".
Последовательность событий, приводящих к активации транскрипции:
- гормон проходит через двойной липидный слой клеточной мембраны. взаимодействие гормона с рецептором (R) приводит к изменению конформации рецептора и снижению сродства к белкам-шаперонам, отделяющимся от комплекса гормон-рецептор. комплекс гормон-рецептор проходит в ядро, взаимодействует с регуляторной нуклеотид-ной последовательностью в ДНК - энхансером или сайленсером. увеличивается (при взаимодействии с энхансером) или уменьшается (при взаимодействии с сайленсером) доступность промотора для РНК-полиМеразы. соответственно увеличивается или уменьшается скорость транскрипции структурных генов. увеличивается или уменьшается скорость трансляции. изменяется количество белков, которые могут влиять на метаболизм и функциональное состояние клетки.
Эффекты гормонов, которые передают сигнал через внутриклеточные рецепторы, нельзя наблюдать сразу, так как на протекание матричных процессов (транскрипцию и трансляцию) требуются часы.
Для исследователей, имеющих представление о количестве сигнальных молекул, о соответствующем количестве рецепторов, о трансмембранных системах передачи сигналов, вторичных посредниках, остаётся загадкой, как протеинкиназы выбирают соответствующий фермент метаболического пути для фосфорилирования. Исследователи для объяснения этого явления предлагают "гипотезу мишени" (от англ, targeting hypothesis). По этой гипотезе специфичность протеинкиназ и фосфопротеинфосфатаз достигается путём образования компартментов на мембране, в состав которых входят не только сами протеинкиназы и фосфопротеинфосфатазы, но и специфические белки-субстраты. Наличие остатка миристиновой или пальмитиновой кислоты в структуре белков-субстратов - условие их "заякоривания" в соответствующем мембранном компартменте.
Специфичность сигнализации
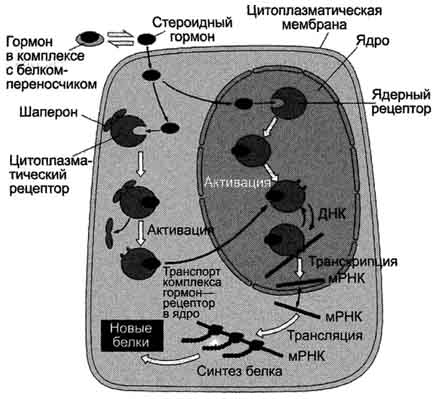
Рис. 5-43. Передача сигнала на внутриклеточные рецепторы.
Арахидоновая кислота и ее продукты: введение
Арахидоновая кислота - ( эйкозатетраеновая кислота ;5,8,11,14-эйкозатетpаеновая) - полиненасыщенная жирная кислота , входит в состав мембранных фосфолипидов тромбоцитов и эндотелиальных клеток . Высвобождается из фосфолипидов под действием фосфолипазы А2 и фосфолипазы С ( рис. 2.3 allerg), активируемых коллагеном , тромбином , АДФ и другими биологически активными веществами. Арахидоновая кислота является предшественником в синтезе оксилипинов см. рис. 1-1 и рис. 1-2 .Свободная арахидоновая кислота быстро метаболизируется, превращаясь в высокоактивные соединения - простагландины и тромбоксаны.
В 90-е годы были получены данные, позволяющие рассматривать AA и ее продукты в качестве еще одной системы вторичных посредников . Во многих случаях показано, что AA и ее производные могут взаимодействовать с другими системами передачи информации в клетке, модулируя их сигналы. Обнаружено, что AA или ее продукты могут влиять на активность фосфолипазы С ( PLC ), аденилатциклазы ( AC ), гуанилатциклазы ( ГЦ ), протеинкиназы С ( PKC ) и приводить к освобождению Са2+ из внутриклеточных депо .
Арахидоновой кислоте приписывается важная роль в регуляции лиганд-рецепторных взаимодействий, активности ионных каналов и активности регуляторных ферментов ( гуанилатциклазы , протеинкиназы C ) в качестве внутриклеточного мессенджера (см. обзор Bonventre, 1992 ). Возможно, что, по крайней мере, некоторые из перечисленных проявлений активности арахидоновой кислоты опосредуются через взаимодействие с клеточными связывающими жирные кислоты белками. Существует два основных пути метаболизма арахидоновой кислоты - циклоксигеназный и липоксигеназный. Циклоксигеназный путь метаболизма арахидоновой кислоты приводит к образованию простагландинов и тромбоксана A2 , липоксигеназный - к образованию лейкотриенов. В тучных клетках легких синтезируются как простагландины, так и лейкотриены, в базофилах - только лейкотриены. Основной фермент липоксигеназного пути метаболизма арахидоновой кислоты в базофилах и тучных клетках - 5-липоксигеназа . 12-липоксигеназа и 15-липоксигеназа играют меньшую роль. Однако образующиеся в незначительном количестве 12-гидропероксиэйкозотетраеновая кислота и 15-гидропероксиэйкозотетраеновая кислота играют важную роль в воспалении .

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
