

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Критерии вида

Существуют признаки, по которым виды отличаются друг от друга. Их называют видовыми критериями (рис.).*
*В литературе можно встретить различные названия и толкования критериев вида: репродуктивный: особи одного вида могут скрещиваться и давать плодовитое потомство; физиолого-биохимический: объединяет два критерия, биохимический и физиологический, исторический: учитывает общее происхождение организмов и др. Ни один из критериев не является абсолютным, видовую принадлежность можно определить лишь по нескольким критериям.
Морфологический | Биохимический | Генетический | |
Самый древний и часто применимый. Основывается на особенностях внешнего строения. С его помощью легко определить виды, которые внешне значительно отличаются. Однако в природе есть большое число видов, практически не отличимых морфологически – видов-двойников (рис.).
Как найти ту границу в морфологических особенностях, которая позволит четко разделить особей на самостоятельные виды? Определить её очень сложно, и видимо, не всегда возможно. Для видового различия можно использовать понятие о хиатусе; хиатус – это разрыв в изменении признака. Если при учете признака обнаруживается перерыв в его значении, то это может служить видовым признаком. Пример, у одном группы особей мушек-дрозофил на бедрах задних конечностей есть от 2 до 6 щетинок, а у другой группы от 9 до 12 щетинок. Есть разрыв в варьировании признака, значит, возможно, это разные виды. | Критерий стал применяться недавно, когда были созданы и распространены новые методы исследования химических соединений, особенно белков и нуклеиновых кислот. Сравнивая нуклеотидный и аминокислотный составы веществ, можно сделать видовой принадлежности особи. Но многие виды имеют внутривидовой биохимический полиморфизм: в пределах популяции существуют различия в аминокислотном составе белков и нуклеотидном составе ДНК. Отсюда разница в морфологических и физиологических характеристиках: уровне кислотности внутренней среды, устойчивости тканей, окраске и др. Пример: у улиток ципей (Cepеa nemoralis) существуют значительные различия в окраске раковины: она может быть желтой, розовой, оранжевой, красной, коричневой. Если использовать морфологический критерий, легко принять разных особей за разные виды. Но это не так; окраска раковины определяется множественным аллелизмом: каждый аллель определяет развитие особой окраски. | Основан на сходстве особей одного вида в организации генетического материала, прежде всего - числа и морфологии хромосом. Все особи одного вида имеют одинаковый набор хромосом (кариотип). Организмы с разным набором хромосом принадлежат к разным видам. Примеры: по числу хромосом отличаются 2 вида–двойника черных крыс. У человека гаплоидный набор хромосом составляет 23, а у высших человекообразных обезьян (шимпанзе, горилла, орангутанг) – 24. При этом одна из крупных хромосом человека соответствует двум мелким хромосомам этих обезьян. Разные виды могут иметь одинаковое число хромосом: зубр и бизон имеют в соматических клетках по 60 хромосом, но гомологичные хромосомы различаются по расположению генов в хромосоме. | |
Физиологический | Этологический | Экологический | Географический |
Заключается в сходстве жизненных процессов, в первую очередь в возможности скрещивания между особями одного вида. Между видами существует физиологическая изоляция. Пример: у многих видов дрозофилы сперма особей чужого вида вызывает иммунологическую реакцию в половых путях самки, что приводит к гибели сперматозоидов. | Связан с разницей в поведении особей, особенно в период спаривания. Особи одного вида имеют очень высокую степень совпадения. Незначительные отличия в ритуале ухаживания, обмене зрительными, звуковыми, световыми сигналами будут препятствовать скрещиванию. Примеры: разница между чайкой полярной и чайкой Тейера в том, что их самцы при ухаживании делают 2 или 3 кивка головой. Эта особенность определяет неспособность чаек к спариванию. Североамериканские светляки обмениваются вспышками света: виды различаются по длине и частоте вспышек. | Основан на том, что каждый вид в пределах своего ареала занимает определенную среду обитания или экологическую нишу и приспособлен к существованию в ней. Особи одного вида обладают одинаковыми биотическими и абиотическими экологическими потребностями. Разные виды в пределах одного ареала характеризуются разными экологическими потребностями. Пример: вьюрки, обитающие совместно на Галапагосских островах, имеют разную пищевую специализацию. | Основан на относительной самостоятельности ареала каждого вида. Разные виды отличаются по площади, размерам, прерывистости ареалов. Пример: хорошо различаются ареалы черной и серой ворон: первый вид обитает на большей части Европы, второй – на Урале, в Сибири и в Западной Европе. Но совпадают ареалы кедрового стланника, даурской лиственницы и душистого тополя, так как эти виды входят в одно растительное сообщество. Есть быстро расширяющие свой ареал виды: растения сорняки, черви-паразиты и др. |
|
|
| |
|
|
|







Трудности в определении вида
Итак, вид – это сложное и неоднозначное понятие. Можно дать такое определение вида: это совокупность популяций с общими морфофизиологическими признаками, способных в природных условиях к скрещиванию друг с другом и биологически изолированных от популяций других видов. В конечном итоге самым важным признаком, отличающим один вид от другого, является не скрещиваемость особей или неспособность их дать нормальное потомство.
Определение применимо к видам, которые размножаются половым путем и не подходит к видам с бесполым, партеногенетическим размножением, а также самооплодотворяющимся. Таких организмов очень много: многие простейшие и черви, грибы и т. д. Данное определение непригодно к вымершим формам. Наконец, его трудно использовать в случаях незавершенного видообразования.
В этих случаях под видом подразумевается группа фенотипически (морфологически) сходных особей, которые занимают сходную экологическую нишу, например, паразитируют в одних и тех же хозяевах. В палеонтологии часто используют не видовые, а родовые названия.
61. Видообразование – микроэволюционный
процесс
Итак, образование нового вида – ключевой момент всего эволюционного процесса. Со времен возникновения жизни на Земле существовало несколько сотен миллионов видов; в настоящее время одних членистоногих существует более миллиона. По Дарвину, в основе видообразования лежат два принципа: монофилии и дивергенции. Рассмотрим пример: группа особей попадает в новые условия или происходит смена одних условий на другие. В этом случае естественный отбор будет отбирать особей с новыми, соответствующими этой среде признаками. Эти признаки будет непрерывно поставлять механизм случайной (ненаправленной) наследственной изменчивости и другие элементарные эволюционные факторы. Даже минимальные, но полезные наследственные признаки будут подхвачены естественным отбором и постепенно разойдутся по всем особям группы. Накапливающиеся различия между «старыми» и «новыми» особями будут столь велики, что через какое-то время можно будет говорить о новом виде. Монофилия, дивергенция, медленное, постепенное накопление небольших наследственных изменений – вот главные принципы дарвинского видообразования.
В последующие десятилетия дарвиновские эволюционные принципы были значительно детализированы и дополнены последователями СТЭ.
Современные данные о микроэволюции

Ключевым моментом для начала микроэволюционных процессов является изоляция. Пока особи разных популяций внутри вида могут скрещиваться и обмениваться наследственным материалом, вид остается целостной и единой системой. Однако в результате изоляции поток генов между популяциями прекращается. Отсутствие потока генов приводит к тому, что популяции генетически дифференцируются: в каждой из них постепенно накапливаются наследственные изменения, полезные для местных условий. Эти наследственные изменения касаются всех сторон жизни особей: поведения, сроков размножения, физиологии, морфологии, пищевой специализации и т. д.
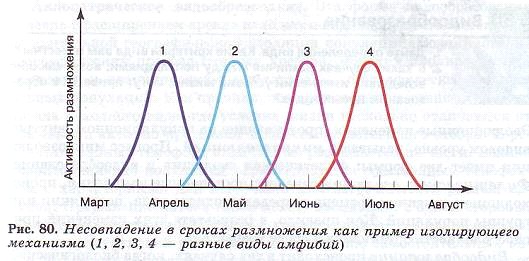
Постепенно формируются репродуктивные изолирующие механизмы (РИМ): комплекс условий, предотвращающих скрещивание между особями изолированных популяций.
Сначала идёт формирование постзиготическихРИМ, когда оплодотворение происходит, но существуют препятствия для образования полноценных гибридов:
1. Гаметический барьер: после слияния гамет образовавшаяся клетка не делится и погибает.
2. Зиготический барьер: зигота образуется, но быстро погибает.
3. На стадии эмбриона или личинки происходит выкидыш плода.
4. Развиваются нежизнеспособные гибриды, которые не справляются с факторами окружающей среды и погибают.
5. Появляются стерильные гибриды, которые не дают собственного потомства.
Эти механизмы предотвращают распространение генетического материала между видами. Но постзиготические механизмы не выгодны: особи вида тратят много времени и энергии на ухаживание, постройку гнёзд, спаривание, но полноценного потомства не получается. Поэтому отбор способствует появлению презиготических РИМ, более выгодных: особи не расходуют силы на ненужные действия. Презиготические РИМ вырабатываются особенно быстро, если изоляция между популяциями исчезает. Есть следующие виды презиготических РИМ:

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 |
