

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Федеральное Государственное Учреждение
«Российский научный центр рентгенорадиологии Федерального агентства по высокотехнологичной
медицинской помощи»
Институт Общей генетики им. РАН
Биологическая индикация радиационного воздействия на организм человека с использованием цитогенетических методов
Медицинская технология №ФС-2007/015-У
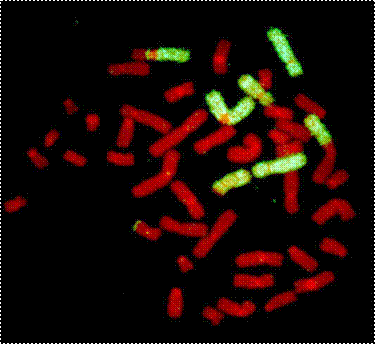
Москва 2007

Аннотация
Представлена усовершенствованная медицинская технология цитогенетического обследования лиц, подвергшихся воздействию ионизирующего излучения в результате профессиональной деятельности или радиационных аварий. Представлены возможности использования цитогенетических методов для оценки уровня облучения людей, как в ранние сроки, так и в отдаленном периоде после радиационного воздействия.
Медицинская технология предназначена для врачей-лаборантов, врачей-генетиков.
Медицинская технология может быть использована в центрах медицины катастроф (федеральных, региональных, территориальных), в диагностических центрах, в центрах государственного санитарно-эпидемиологического надзора в субъектах Российской Федерации и Федерального управления медико-биологических и экстремальных проблем Минздрава России.
Учреждение ФГУ «Российский научный центр рентгенорадиологии
разработчик Росмедтехнологий», ГСП-7, Москва, ул. Профсоюзная, д.86
Соразработчик: Институт Общей генетики им. РАН
ГСП-1, Москва, ул. Губкина, д.3
Разработчики:
- ведущий научный сотрудник отдела радиационной медицины РНЦРР,
кандидат биологических наук
- биолог отдела радиационной медицины РНЦРР, кандидат биологичес-
ких наук
- инженер-химик отдела радиационной медицины РНЦРР
- инженер-программист отдела радиационной медицины РНЦРР
- заведующий лабораторией радиационной генетики ИОГен РАН,
доктор биологических наук
Рецензенты:
- руководитель кариологической лаборатории ФГУ ГНЦ РАМН,
доктор медицинских наук, профессор;
- руководитель лаборатории радиобиологии ФГУ ГНЦ РФ – ИМБП РАН, доктор биологических наук
Введение
В профессиональных условиях работы с источниками ионизирующих излучений возможно возникновение нештатных ситуаций, когда определение поглощенной дозы с помощью физической дозиметрии может быть ограничено или невозможно. В этих случаях могут быть применены биологические методы дозиметрии, которые позволяют оценить суммарный эффект от воздействия радиации с учетом индивидуальных особенностей облученного организма. Такая информация имеет, несомненно, большое значение для оказания своевременной эффективной помощи в случае острого радиационного воздействия, а также для прогностической оценки возможных отдаленных последствий облучения.
Наибольшее распространение получили методы биодозиметрии, основанные на анализе частоты радиационно-индуцированных генетических изменений в соматических клетках – геномных, хромосомных или генных мутаций. Одним из самых разработанных и обоснованных является цитогенетический метод, при котором в качестве биологического маркера облучения используются хромосомные аберрации в лимфоцитах периферической крови. Материалы многочисленных отечественных и зарубежных цитогенетических исследований послужили основой для выработки рекомендаций ВОЗ, МАГАТЭ, НКДАР ООН по практическому использованию данного метода при определении доз облучения (WHO, 1976; UNSCEAR, 1982; IAEA, 1986).
Радиобиологической основой для применения цитогенетического метода в дозиметрии является высокая чувствительность лимфоцитов периферической крови, а также наличие специфичных для радиации хромосомных перестроек, дозовая зависимость частоты которых для большинства видов ионизирующих излучений хорошо изучена. Под воздействием радиации могут возникать два типа хромосомных аберраций: нестабильные (дицентрики, центрические кольца, ацентрические фрагменты) и стабильные (реципрокные и другие типы транслокаций). Чаще всего для оценки величины поглощенной организмом дозы используют частоту возникновения обменных аберраций нестабильного типа, а именно, дицентрических хромосом. Преимущество дицентриков состоит в том, что аберрации данного типа являются специфичными для воздействия ионизирующих излучений, их легко обнаружить в световом микроскопе без применения сложных методов обработки и окрашивания хромосомных препаратов. Однако следует иметь в виду, что клетки, несущие такие аберрации, образуют при делении генетически несбалансированные клетки, что в дальнейшем приводит к их элиминации. Поэтому успешное применение нестабильных аберраций для определения дозы облучения в основном возможно в ранние (до 3 - 4 месяцев) сроки после однократного, относительно равномерного радиационного воздействия. В случае хронического облучения и при ретроспективной оценке дозы радиационного воздействия необходимо применение различных математических моделей, учитывающих либо процесс элиминации подобных клеток из кровотока, либо особенности распределения аберраций по клеткам
Определить степень радиационного поражения организма, а также оценить дозу облучения спустя длительное время после радиационного воздействия или в случае хронического облучения можно с помощью стабильных хромосомных аберраций (транслокаций) в лимфоцитах периферической крови. Это возможно благодаря тому, что такие повреждения хромосом не нарушают течения митоза и могут передаваться в ряду клеточных поколений, то есть частота транслокаций не должна подвергаться значительным изменениям со временем.
В последние годы для определения частоты стабильных хромосомных аберраций (транслокаций) широко используют новый молекулярно-генетический метод – FISH (fluorescence in situ hybridization), который позволяет визуализировать отдельные хромосомы или участки хромосом. Он основан на флуоресцентном окрашивании хромосом или отдельных участков хромосом после гибридизации in situ с использованием хромосомо-специфичных ДНК-зондов. В препаратах метафазных хромосом, окрашенных с помощью FISH метода, структурные нарушения легко распознаются, поэтому скорость анализа препаратов значительно выше, чем при использовании дифференциального окрашивания, и, кроме того, не требуется долгое обучение исследователя.
Показания к использованию медицинской технологии
Возможное радиационное воздействие на организм человека в результате различных чрезвычайных и аварийных ситуаций.
Противопоказания к использованию медицинской технологии
Противопоказаний к применению метода нет.
Материально-техническое обеспечение медицинской технологии
Для осуществления медицинской технологии используется стандартный комплект оборудования и реактивов для проведения цитогенетического обследования, согласно приказу МЗ РФ № 000 от 01.01.2001 г.
Описание медицинской технологии
Цитогенетическое исследование включает следующие основные стадии:
1. Получение препаратов метафазных хромосом из лимфоцитов периферической крови.
2. Окрашивание препаратов.
3. Микроскопический анализ и статистическая обработка результатов.
Получение препаратов хромосом из лимфоцитов периферической крови
В заранее приготовленные стерильные пробирки с гепарином ед. гепарината лития в расчете на 1 мл крови) вносят 5 - 6 мл крови и закрывают стерильной пробкой. Для постановки культуры обычно используют цельную кровь, можно также использовать плазму, обогащенную лейкоцитами. Для этого применяют осторожное центрифугирование крови (800-900 об/мин) в течениемин, после чего стерильной пипеткой собирают пленку лейкоцитов, располагающуюся на границе эритроцитов с плазмой. Наилучшие результаты получаются при использовании свежей крови, тем не менее, можно получить удовлетворительный митотический индекс и после хранения стерильной крови при комнатной температуре в течение нескольких дней, при этом лучше хранить плазму с лейкоцитами.
· Приготовление культуральной среды
Описанные ниже процедуры проводят, строго соблюдая условия стерильности.
Состав культуральной среды:
§ Питательная среда (типа RPMI-1640, Игла или 199);
§ 15-17% сыворотки (крупного рогатого скота, телячьей или эмбриональной телячьей);
§ 2 мМ глутамина;
§ антибиотики - пенициллин (100 МЕ/мл), стрептомицин (0,1 мкг/мл).
Стерильным шприцем или пипеткой разливают по 4.5 мл культуральной среды во флаконы и закрывают крышками. В таком виде среда для культивирования при необходимости может храниться при t = -20оС в течение месяца.
Во флаконы с культуральной средой (4,5 мл) стерильной пипеткой вносят по 0,5 мл крови или плазмы, обогащенной лимфоцитами, добавляют 5-бромдезиксиуридин (БДУ) в
конечной концентрации 3-6 мкг/мл, 2 – 2,5% фитогемагглютинина и помещают в термостат при t=37оС.
Для накопления клеток на стадии метафазы митоза используют ингибиторы образования веретена деления – колхицин или колцемид. Конечная концентрация колхицина в культуральной среде должна быть не более 0,25 – 0,40 мкг/мл, колцемида – 0,1–0,2 мкг/мл. Такие количества блокирующего вещества позволяют получить достаточное количество метафаз без заметного токсического эффекта, который может иметь место при более высоких концентрациях. Удобным для использования является раствор колцемида в физиологическом растворе с концентрацией 5 мкг/мл. Стерильно приготовленный раствор колцемида можно хранить при t=4оС в течение нескольких месяцев. Через 45 часов от начала культивирования 0.15 мл такого раствора добавляют в каждый флакон, культуру клеток перемешивают и оставляют в термостате при t=37оС на 3 часа.
По окончании инкубации с колцемидом (или колхицином) тщательно перемешивают содержимое флаконов и переливают в 15 мл центрифужные пробирки. Дальнейшая обработка клеточной суспензии проходит при комнатной температуре. Пробирки центрифугируют при 1200 об/мин в течение 5-8 мин для осаждения клеток. Затем пастеровской пипеткой или при помощи водоструйного насоса отбирают супернатант, оставляя около 0,5 мл надосадочной жидкости. Осадок из клеток ресуспендируют в 8-10 мл гипотонического раствора, предварительно нагретого до 37оС. В качестве гипотонического раствора можно использовать 0,56% раствор хлористого калия; раствор, содержащий равные количества 0,56% раствора хлористого калия и 0,95% раствора цитрата натрия; раствор Хенкса, разбавленный дистиллированной водой в объемном соотношении 1 к 4. Пробирки с гипотоническим раствором на 15 мин помещают в термостат при t=37оС, затем их снова центрифугируют при 1000 об/мин в течение 5 мин, удаляют гипотонический раствор, оставляя 0,5 мл супернатанта. Общее время гипотонической обработки должно быть примерно 20-25 мин. Точная продолжительность гипотонической обработки подбирается опытным путем.
После гипотонии к клеточному осадку добавляют свежеприготовленный холодный фиксатор (смесь метилового спирта с ледяной уксусной кислотой в объемном соотношении 3 к 1). Фиксатор добавляют медленно, равномерной струйкой, постоянно перемешивая при помощи вихревого смесителя или пастеровской пипетки для предотвращения образования комков или агрегатов клеток. Через несколько минут (за это время происходит полное разрушение эритроцитов) клетки осаждают центрифугированием при 1000 об/мин в течение 5 мин, отбирают супернатант и добавляют к осадку 5 - 7 мл свежего фиксатора. Эту процедуру повторяют 3 - 4 раза, при этом полная продолжительность фиксации клеток должна составлять не менеемин. Замену фиксатора прекращают, когда клеточный осадок приобретает белый цвет. При последней промывке удаляют супернатант, оставляя примерно 0,25 – 0,5 мл клеточной суспензии.
· Приготовление препаратов хромосом
Одним из условий для получения качественных препаратов хромосом является подготовка предметных стекол. Поверхность стекла должна быть идеально чистой! Клетки тщательно ресуспендируют в оставшемся фиксаторе с помощью пастеровской пипетки, наносят 3 капли клеточной суспензии с высоты около 10 см на предметное стекло и высушивают его на термоплате при t = 45 – 48оС. Качество препарата оценивают под микроскопом. Хороший препарат должен содержать при малом увеличении (´200) в поле зрения не менее 1 - 2 метафазных пластин с расправленными хромосомами без взаимных наложений. При необходимости, если клеток очень много, можно разбавить суспензию фиксатором или наоборот, если их мало, можно осадить клетки центрифугированием при 1000 об/мин в течение 5 мин и затем отобрать некоторое количество супернатанта.
Препараты метафазных хромосом, предназначенные для анализа нестабильных хромосомных аберраций, перед окрашиванием выдерживают в темноте при комнатной температуре в течение 3-5 дней. Препараты метафазных хромосом, которые предназначены для анализа стабильных хромосомных аберраций, рекомендуется использовать для окрашивания в течение первых двух недель после приготовления, оптимальным сроком хранения считается неделя. Для более длительного хранения препараты метафазных хромосом помещают в герметично запаянные полиэтиленовые пакеты, заполненные газообразным азотом, и хранят в холодильнике при –20оС. Возможно также хранение клеток в виде фиксированной клеточной суспензии при –20оС.
Анализ нестабильных хромосомных аберраций
Окрашивание методом «флуоресценция плюс краситель Гимза» (FPG метод)
1. Поместить стекла в ванночку с раствором красителя Hoechst 33258 в PBS (конечная концентрация 0,25 мкг/мл) и выдерживать при комнатной температуре в темноте в течение 15 мин.
2. Поместить ванночку со стеклами на термоплату (t = 60оС) под ультрафиолетовую лампу (265 нм) на 20 мин.
3. Вынуть стекла из раствора красителя, промыть проточной дистиллированной водой.
4. Опустить стекла на 5 мин в 5% раствор красителя Гимза в фосфатном буфере (pH 6,8).
5. Промыть стекла в дистиллированной воде и высушить на воздухе.
Препараты просматривают в световом микроскопе при малом увеличении (´160 или ´200) и выбирают для анализа хорошо окрашенные метафазные пластинки, без наложений хромосом. Затем при увеличении ´1000 под масляной иммерсией анализируют только метафазы I митоза, хромосомы которых равномерно окрашены (метафазы II митоза имеют «арлекинную» окраску) и имеют в сумме 46 центромер. Учитывают все типы хромосомных аберраций и заносят их в специальный протокол. При биодозиметрических исследованиях особое внимание уделяют дицентрическим и кольцевым хромосомам, которые являются маркерами радиационного воздействия.
Анализ стабильных хромосомных аберраций
Для обнаружения транслокаций применяют метод окрашивания, основанный на гибридизации in situ с использованием ДНК проб, специфичных к отдельным хромосомам человека, т. н. FISH метод. Метод FISH основан на способности хромосомной ДНК (ДНК-мишень) связываться в определенных условиях с фрагментами меченной ДНК (ДНК-зонд), включающими нуклеотидные последовательности, комплементарные ДНК-мишени.
Ниже описана модифицированная методика гибридизации in situ Пинкеля (Pinkel et al.,1986) с использованием в качестве зондов биотинилированных ДНК_проб, специфичных к отдельным хромосомам человека, а также панцентромерных ДНК-проб, меченных дигоксигенином.
Можно использовать готовые наборы для анализа транслокаций фирмы MetaSystems (http://www. metasystems. de). При работе следует руководствоваться инструкцией, прилагаемой к набору.
· Предварительная обработка препаратов метафазных хромосом
Хорошее качество препаратов метафазных хромосом является залогом успешной гибридизации in situ. Важно, чтобы хромосомы на стекле располагались относительно свободно, без взаимных наложений, чтобы количество окружающей их цитоплазмы было минимальным. При большой плотности клеточного материала на стекле реакция гибридизации может быть неэффективной и /или может наблюдаться интенсивный фон вследствие неспецифического связывания ДНК-зонда. Существуют разные способы предварительной обработки препаратов, позволяющие избавиться от нежелательных эффектов. Ниже приведены описания процедур, с помощью которых можно улучшить качество метафазных препаратов.
· Обработка пепсином
Предварительная обработка препаратов пепсином позволяет в некоторых случаях избавиться от нежелательного фона. Данная процедура не является обязательной.
Поместить сосуд Коплина со 100 мл дистиллированной воды в водяную баню при t=37оС. За 15 мин до работы со стеклом в воду добавить 100 мкл 10 N HCl, за 10 мин до обработки пепсином добавить 50 мкл 10% пепсина.
1. Поместить стекло на 2 мин в PBS при комнатной температуре.
2. Поместить стекло в 10% раствор пепсина на 10 мин, t= 37оС.
3. Перенести стекло в сосуд Коплина с PBS на 2 мин при комнатной температуре.
4. Промыть стекло в течение 2 мин в растворе PBS/MgCl2 при комнатной температуре на шейкере.
· Обработка параформальдегидом
Обработка параформальдегидом позволяет лучше сохранить структуру хромосом в процессе их окрашивания. Эта процедура также не является обязательной.
1. Погрузить стекло на 1 мин при комнатной температуре в 0,25% раствор параформальдегида.
2. Промыть стекло в течение 2 мин в PBS при комнатной температуре. Использовать шейкер.
3. Провести стекло через серию спиртов - 70%, 90% и 100% растворы по 2 мин при комнатной температуре.
4. Просушить стекла на воздухе или струей газообразного азота.
· Денатурация и гибридизация.
Необходимо заранее приготовить 70% раствор формамид/2*SSC в сосуде Коплина и нагреть его до 72оС на водяной бане.
Обработанное с помощью параформальдегида стекло помещают в 70% раствор формамида на 4 мин при t=72оС. Для стекол, не прошедших обработку параформальдегидом, время денатурации сокращается до 2 мин.
Из 70% раствора формамида стекла перенести в охлажденный до t=-20оС 70% этанол на 2 мин, далее инкубировать по 2 мин в 90% и 100% этаноле при комнатной температуре. Просушить стекла на воздухе или струей газообразного азота.
Поместить стекла на термоплату, t=37оС, нанести гибридизационную смесь, покрыть покровным стеклом так, чтобы под ним не было пузырей воздуха и заклеить края покровного стекла резиновым клеем. Дождаться, пока клей высохнет, положить стекла в предварительно прогретую влажную камеру и поместить ее в термостат t=37оС. Для успешной гибридизации достаточно 12 – 18 часов, в случае необходимости можно продлить срок гибридизации до 3 суток.
· Состав и приготовление гибридизационной смеси
При приготовлении гибридизационной смеси необходимо работать только в перчатках, желательно проавтоклавировать используемую одноразовую пластиковую посуду и бидистиллированную воду при 1 атм 120 мин.
Гибридизационную смесь с хромосомными пробами для одного стекла готовят в одной микропробирке (конечный объем гибридизационной смеси для 3-х хромосом – 30 мкл). Гибридизационную смесь с панцентромерной пробой готовят в отдельной микропробирке. Перед нанесением на стекло содержимое микропробирок с хромосомными пробами и панцентромерной пробой объединяют. Когда качество проб неизвестно и требуется определить наиболее оптимальные для конкретных проб концентрации, используется стандартный состав проб (Таблица 1). В случае недостаточно хорошего окрашивания хромосом-мишеней при стандартном составе гибридизационной смеси следует увеличить количества проб тех хромосом, которые плохо окрашиваются, при этом следует, соответственно, уменьшить количество добавляемой бидистиллированной воды (вплоть до 0 мкл) и/или Cot-1 ДНК (общее конечное количество можно уменьшить до 4 мкл). Количество Master Mix 1.0 и конечный объем гибридизационной смеси менять нельзя!
Таблица 1. Стандартный состав гибридизационной смеси для окрашивания хромосом по методу FISH с использованием ДНК-проб для хромосом 1, 4 и 12 и панцентромерных проб
Исходные реагенты | #1 * мкл | #4 * мкл | #12 * мкл | Панцентромерная проба **, мкл |
ДНК-проба (»150 мкг/мл) | 1,0 | 1,0 | 1,0 | 3,0 |
Master Mix 1,0 | 7,0 | 7,0 | 7,0 | 7,0 |
Cot-1 ДНК человека (1 мг/мл) | 2,0 | 2,0 | 1,0 | |
H20 бидистилл. | - | - | 1,0 | |
Конечный объем | 10,0 | 10,0 | 10,0 | 10,0 |
* Биотинилированные ДНК пробы, специфичные к хромосомам 1, 4 и 12, получают при помощи ник-трансляции Hind III библиотек pBS-1, pBS-4 и pBS-12. Концентрация ДНК-проб варьирует от 150 до 200 мкг/мл.
** Меченные при помощи digoxigenin-dUTP панцентромерные пробы получают с использованием полимеразной цепной реакции. Концентрация панцентромерных проб варьирует от 100 до 250 мкг/мл.
1. Пробирки с хромосомными пробами, Cot-1 ДНК и Master Mix 1.0 достать из холодильника. Разморозить, отцентрифугировать на миницентрифуге в течение нескольких секунд, затем желательно поставить в лед.
2. Компоненты гибридизационной смеси с помощью автоматической микропипетки перенести в микропробирку (каждый компонент своим носиком, носик не использовать вторично даже в случае, когда отбирается один и тот же компонент смеси). Отцентрифугировать на микроцентрифуге в течение 2 - 3 сек, перемешать на шейкере и снова отцентрифугировать.
3. Поместить микропробирку с гибридизационной смесью на 5 мин в водяную баню, t = 72оС.
4. Поместить микропробирку с гибридизационной смесью для быстрого охлаждения на лед на несколько секунд.
5. Микропробирку с гибридизационной смесью и хромосомными пробами поместить в термостат (37оС) на 30 – 60 мин.
6. Денатурация панцентромерной ДНК-пробы также проводится на водяной бане при t = 72оС в течение 5 мин. и затем переносится в лед для быстрого охлаждения на 1 – 5 мин.
7. Содержимое двух микропробирок (с гибридизационной смесью, хромосомными пробами и с панцентромерной пробой) смешать непосредственно перед нанесением гибридизационного коктейля на стекло.
8. Для этого следует отцентрифугировать микропробирку с панцентромерной пробой на микроцентрифуге, перенести ее содержимое в микропробирку с хромосомными пробами и гибридизационной смесью, отцентрифугировать, тщательно перемешать на шейкере и снова отцентрифугировать.
9. Нанести гибридизационный коктейль на препарат, покрыть покровным стеклом (24*50 мм2), заклеить края покровного стекла резиновым клеем. После засыхания клея препарат поместить в предварительно прогретую влажную камеру, и поместить на ночь в термостат, t = 37оС.
· Послегибридизационные отмывки и флуоресцентное окрашивание.
1. Поместить 4 сосуда Коплина с 50% раствором формамида в водяную баню, t=42оC. Приготовить 300 мл раствора 0,1*SSC, разлить по трем сосудам Коплина, поместить их в водяную баню, t = 60оC.
2. Снять пинцетом резиновый клей со стекла. Для того, чтобы смыть покровное стекло, препарат погрузить на 5 мин в 50% раствор формамида, t=42оC. После удаления покровного стекла препарат должен оставаться в первом растворе формамида 5 мин.
3. Повторить трижды по 5 мин отмывку в 50% растворе формамида, t= 42оC.
4. Трижды по 5 мин отмывать в 0,1*SSC при t=60оC. При проведении окрашивания на вторые митозы на этой стадии удобно провести обработку стекол раствором Hoeсhst (как описано выше).
5. Поместить стекло в PN-буфер на 5 мин при комнатной температуре.
6. Нанести на стекло 100 мкл PNM буфера, покрыть пленкой из плотного полиэтилена и поместить стекло во влажную камеру. Поставить влажную камеру в термостат t=37оC на 20 мин.
7. После обработки PNM буфером пленку аккуратно снять, препарат поставить вертикально, чтобы стекли остатки PNM буфера, при этом стекло не должно высыхать.
8. На препарат нанести 100 мкл раствора, содержащего равные количества растворов антидигоксигенина и стрептавидина-FITC, покрыть полиэтиленовой пленкой, поместить стекло во влажную камеру. Окрашивание происходит в темноте в течение 20 мин при комнатной температуре.
9. Промыть дважды по 2 мин в PN-буфере. Использованный буфер выливается.
10. На препарат нанести 100 мкл раствора, содержащего равные количества растворов биотинилированного антистрептавидина и антител IgG Rat-anti-Mouse-AMCA, покрыть полиэтиленовым покровным стеклом, поместить стекло во влажную камеру, окрашивание вести в темноте в течение 20 мин при комнатной температуре.
11. Промыть дважды по 2 мин в PN-буфере.
12. На препарат нанести 100 мкл раствора, содержащего равные количества растворов стрептавидина-FITC и антител IgG Mouse-anti-Rat-AMCA, покрыть полиэтиленовым покровным стеклом, поместить стекло во влажную камеру, окрашивание вести в темноте в течение 20 мин при комнатной температуре.
13. Стекло промыть дважды по 2 мин в PN-буфере.
14. Поставить стекло на несколько секунд вертикально для того, чтобы стекли излишки PN-буфера. Нанести на стекло 22 мкл раствора пропидия иодида в Antifade (PI), покрыть покровным стеклом (24*50мм2), под микроскопом оценить качество окраски. Если требуется дополнительное окрашивание, то следует аккуратно смыть покровное стекло в PN-буфере и повторить последовательно стадии 10, 11, 12, 13. Готовые окрашенные препараты хранят при 4оС. Если в процессе микроскопирования и/или хранения выцвел FITC, то можно «освежить» окраску, вновь проделав стадии 10, 11, 12, 13 флуоресцентного окрашивания.
· Микроскопический анализ хромосомных аберраций
Готовые препараты анализируют на флуоресцентном микроскопе с помощью набора фильтров, один из которых позволяет наблюдать одновременную флуоресценцию FITC и PI, второй - АМСА-флуоресценцию, третий фильтр обеспечивает флуоресценцию только PI.
При использовании ДНК-проб для хромосом 1, 4, 12 и флуорохромов FITC, PI и AMCA две пары хромосом (4 и 12) полностью окрашиваются в желтый цвет. Хромосомы 1 окрашены в желтый цвет, исключая небольшую дистальную часть короткого плеча и околоцентромерную область, которые окрашены в красный цвет, так же как и другие хромосомы после окрашивания пропидий иодидом. Центромеры после гибридизации с панцентромерными пробами окрашены в голубой цвет. Если все шесть окрашенных хромосом в клетке являются интактными, клетку признают «нормальной». При наличии в клетке аберрации с участием окрашенной желтой хромосомы, по возможности, учитывают, какая именно хромосома участвовала в данной аберрации. Конфигурацию, при которой в клетке имеются две двуцветные хромосомы, взаимно обменявшиеся своими частями, расценивают как «полную» транслокацию Если в клетке одна двуцветная хромосома, а все остальные 45 хромосом красные, транслокацию считают «неполной». Классификация дицентриков, центрических колец и ацентрических фрагментов принципиально не отличается от принятой в рутинном анализе.
Для анализа частоты транслокаций FISH методом могут быть использованы ДНК-зонды для самых разных комбинаций хромосом:1, 4 и 12; 1, 4 и 8; 1, 2 и 4 и т. д. Для того, чтобы сравнивать результаты таких исследований удобно использовать значения геномной частоты транслокаций (FG) и клеточного эквивалента (Neq). Для их определения используют следующие формулы (Lucаs et al., 1992):
FG=FP/2,05*fP*(1-fP),
Neq=2,05*fP*(1-fP)*N,
где FP – наблюдаемая частота транслокаций с участием «окрашенных» хромосом;
N – число проанализированных метафаз, а fP- доля генома, которую составляют «окрашенные» хромосомы (т. е. относительное содержание ДНК «окрашенных» хромосом). Для хромосом 1, 4 и 12 эта величина составляет 0,19 (Mendelson et al., 1973).
Реконструкция дозовой нагрузки по частоте хромосомных аберраций
Для оценки доз в случае острого радиационного воздействия в ранние сроки после облучения проводят анализ нестабильных хромосомных аберраций в лимфоцитах периферической крови классическим цитогенетическим методом. При ретроспективном обследовании, а также в случаях хронического или пролонгированного облучения необходимо проводить анализ стабильных хромосомных аберраций в лимфоцитах крови FISH методом. При реконструкции дозовой нагрузки в первую очередь необходимо оценить уровень значимости различий между спонтанной частотой аберраций и частотой аберраций у облученного пациента (группы пациентов). Если различия являются статистически значимыми, то оценивают дозу облучения с использованием калибровочных кривых «доза-эффект».
Определение статистической значимости наблюдаемых различий частот аберраций
Как уже указывалось выше, статистический анализ цитогенетических данных следует начинать с выяснения вопроса о достоверности повышения частоты аберраций у облученного пациента (группы). Эта стандартная процедура в случае обработки результатов цитогенетического анализа имеет ряд специфических особенностей, что связано с трудностями, возникающими при сравнении частот редких событий. Следует отметить, что многочисленные дискуссии вокруг методов биодозиметрии и радиобиологии малых доз в большинстве случаев связаны со статистической обработкой полученных результатов. При сравнении частот хромосомных перестроек наиболее адекватно использование точного критерия Фишера, поскольку стандартные параметрические критерии (различные модификации t-критерия Стьюдента) являются слишком консервативными в данной ситуации, а для применения непараметрического теста c2 необходимо выявить не менее 5 аберраций как в опыте, так и в контроле. Суть точного критерия Фишера заключается в точном вычислении вероятности Р в предположении, что облученная и контрольная группы неразличимы по частоте аберраций. При анализе цитогенетических данных число проанализированных клеток Ni значительно превышает число выявленных аберраций ni (Ni>>ni). В этом случае значение Р приблизительно равно:

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
