

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Среди животных с ДHF% высок процент симпатотоников - 36%, а ваготоников – только 24%; среди крыс с ДLF%, ДVLF% преобладают особи с В и ВБ (табл. 5). Анализ типов ВСР с учетом половой принадлежности особей установил, что 46% самцов имеют ВБ, 28% - тип В и 26% - тип С. Среди самок большинство - 50% - симпатикотоники, 32% - с ВБ и только 18% - ваготоники. Среди крысят типы ВС и НВБ встречаются в примерно равном соотношении. Для описания типа регуляции СР предложена формула, включающая 1) тип активности уровней регуляции (ДHF%, ДLF% или ДVLF%), 2) тип симпато-парасимпатических отношений (ВБ, С или В у половозрелых животных, ВС или НВБ среди 6-недельных крысят). Соотношение типов ВСР характеризуется половыми и возрастными особенностями, однако во всех случаях среди животных в условиях фоновой активности преобладают особи с автономным типом регуляции: среди самцов особи с ДHF%+ВБ и ДHF%+В, среди самок - ДHF%+С, а среди крысят – с ДHF%+ВС.
У самцов крыс с ДHF%+С активность ХЭ плазмы и ОМН выше, чем у особей с ВБ и В (р<0,05 и р<0,01) (табл. 6). У самок и крысят типологические особенности этих показателей не выражены. Половозрелые самцы с типом С отличаются от особей с ВБ низким уровнем ПОЛ и каталазной активности тканей (р<0,01 и р<0,05), более высокой каталазной активностью плазмы крови и АОА миокарда (р<0,05). Среди самок особи с типом В характеризуются низким уровнем ТБК-РП (р<0,01), высокой каталазной активностью и общей АОА плазмы крови и миокарда (р<0,05). У крысят с ВС уровень ПОЛ в тканях и плазме крови выше, чем у особей с НВБ (р<0,05). Следовательно, с возрастом у крыс с преобладанием симпатических влияний интенсивность ПОЛ в тканях становится ниже, чем у животных с другими типами регуляции СР, то есть возрастное становление вегетативной регуляции функций сопровождается изменением про - и антиоксидантного статуса организма.
Таблица 6
Относительная масса надпочечников, активность холинэстеразы плазмы крови и показатели свободнорадикального гомеостаза нелинейных крыс с различными типами регуляции сердечного ритма (М±m)
Показа-тели | Самцы | Самки | ||||
С (8) | ВБ (10) | В (7) | С (10) | ВБ (7) | В (4) | |
ОМН, мг/г | 0,156± 0,005 # | 0,136± 0,007 | 0,128± 0,005 && | 0,191± 0,018 □ | 0,228± 0,018 □□□ | 0,213± 0,024 □□□ |
ХЭ, ед. опт. пл. | 51,7±2,42 # | 41,2±2,17 | 44,7±1,44 | 113,9±9,5 □□□ | 112±10,1 □□□ | 101±14,1 □□ |
ТБК-РП пл | 2,61±0,28 | 3,17±0,29 | 2,78±0,17 | 2,31±0,15 | 2,53±0,30 | 1,33±0,10 #, &&, □□ |
Катал пл. | 61,3±3,4 # | 46,4±4,1 | 44,7±4,2 && | 47,1±5,8 | 48,7±9,7 | 77,7±1,9 &, □□□ |
АОА пл | 70,3±3,49 | 75,9±4,06 | 66,2±3,58 ## | 80,4±2,29 □ | 83,8±1,29 □ | 88,8±1,44 #, &, □□□ |
ТБК-РП миок | 0,58±0,08 ## | 1,03±0,09 | 0,76±0,07 #, & | 0,32±0,03 □□ | 0,31±0,04 □□□ | 0,33±0,04 □□ |
Катал миок | 14,7±0,6 # | 16,7±0,7 | 16,3±1,5 | 6,8±0,56 ###, □□□ | 10,7±0,6 □□□ | 8,3±0,47 #, □□□ |
АОА миок | 79,9±1,14 # | 71,7±2,76 | 70,1±1,23 &&& | 73,2±2,96 | 75,1±1,88 | 86,4±2,87 #, □□□ |
ТБК-РП печ | 0,81±0,09 ## | 2,03±0,22 | 1,03±0,26 # | 0,55±0,11 | 0,39±0,04 □□□ | 0,29±0,05□ |
Катал печ | 237,9±4,1 ## | 264,1±5,2 | 244,0±5,1 # | 217,±10,0 | 232,3±7,9 □□ | 256,4±7,5 |
Примечание. p<0.05, p<0.01, p<0.001 - #, ##, ### - по сравнению с группой ВБ; &, &&, &&& - по сравнению с группой С, □, □□, □□□ – по сравнению с самцами; ^, ^^, ^^^ - по сравнению с показателями половозрелых крыс соответствующих типологических групп
Проблема стресса в современном обществе является очень острой, поэтому исследования в области физиологии стресса остаются неизменно актуальными. Согласно нашим данным, характерными стресс-индуцированными изменениями у половозрелых крыс и крысят являются сильная тахикардия за счет снижения Мо; снижение мощности HF-волн; рост мощности и доли в спектре волн LF, а также VLF (у самок); повышение IC, которое обнаруживается чаще, чем рост ИН. У крыс всех групп с ростом ЧСР коррелирует повышение каталазной активности плазмы крови (r=0,60 – 0,89, р<0,01) и рост уровня ТБК-РП и скорости АЗ-ПОЛ в печени (r=0,72-0,73 и r= 0,87-0,85, р<0,01).
Выраженность и направленность стресс-индуцированных изменений показателей зависят от возраста, пола и типологических особенностей организма. Так, у крысят абсолютные значения ЧСР и ИН всегда выше, абсолютные мощности волн СР ниже, но степень стрессорных изменений всегда меньше, чем у половозрелых крыс. Крысята, имеющие в покое более ригидный СР, на начальном этапе стресса часто дают повышение вариабельности СР за счет LF и VLF-волн, которое сменяется резким усилением ригидности СР. В свою очередь, у самок, по сравнению с самцами степень прироста ЧСР обычно меньше (около 25% против 37-41%), вариабельность СР за счет LF - и VLF-волн повышается быстрее и сильнее, что приводит к росту IC, нежели к росту ИН, интенсивность ПОЛ и каталазная активность плазмы крови изменяются в меньшей мере.
В зависимости от типа регуляции СР среди крыс с ДHF% в обеих возрастных группах наблюдаются различные варианты (типы) реакции на стресс. При описании этих типов нами применены термины «прессорный» и «депрессорный», под которым понимается повышение или снижение ригидности СР, усиление симпатических или парасимпатических влияний. У половозрелых крыс (рис. 1) с ВБ выявлен сбалансированный прессорно-депрессорный тип реакции с быстрым восстановлением вегетативного баланса на сегментарном уровне, умеренной (у самцов) и сильной (у самок) активацией структур центрального контура регуляции, умеренной интенсификацией ПОЛ в тканях (р<0,05 и р<0,01) и 2-3 кратным ростом каталазной активности плазмы крови (р<0,001). У крыс с типом С реакция на стресс носит взрывной прессорно-депрессорный характер, с резкой интенсификацией ПОЛ (р<0,001) и 3,5-4,5-кратным ростом каталазной активности плазмы крови (р<0,001). Тип реакции на стресс крыс-ваготоников (особенно самцов) можно определить как замедленный (депрессорно-прессорный у самцов, прессорно-депрессорный у самок), так как напряжение СР развивается медленно и сопряжено с сильной активацией структур центрального контура регуляции, значительным накоплением продуктов ПОЛ, 3-1,5-кратным ростом каталазной активности плазмы крови (р<0,001) и снижением устойчивости тканей самок к прооксидантной провокации (р<0,001).
У крысят-самцов с НВБ (рис. 2) реакцию на стресс можно определить как сбалансированную, прессорно-депрессорного типа с умеренной централизацией управления (р<0,01), умеренной активацией ПОЛ (р<0,01) и каталазной активности плазмы (в 2,1-2,4 раза, р<0,001). Крысята с ВС и крысята-самки с НВБ дают на стресс взрывную реакцию депрессорно-прессорного типа с максимальным нарастанием вариабельности СР (р<0,001) в первой половине ЭБС, резким повышением каталазной активности плазмы крови (в 2,4-3,4 раза, р<0,001) и выраженной активация ПОЛ в тканях (р<0,001 - р<0,05).
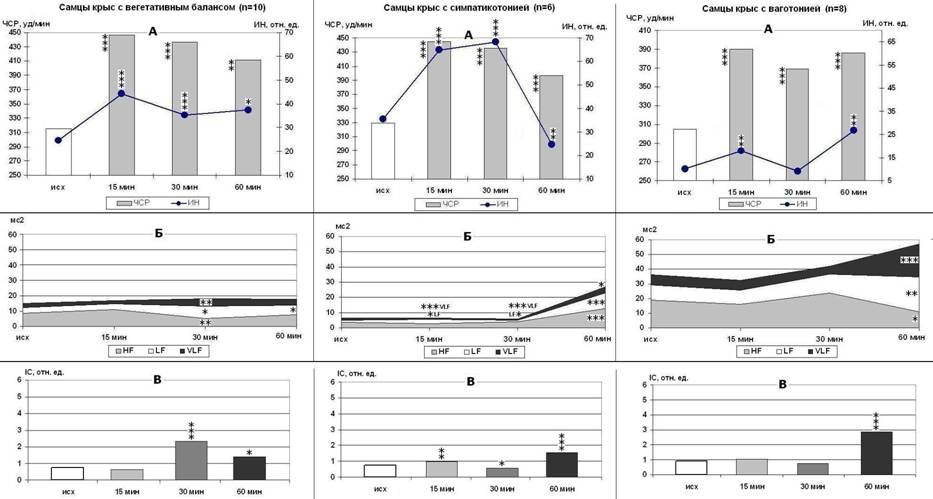
Рисунок 1. Изменения ЧСР и ИН (А), мощности волн спектра ВСР (Б) и IC (В) у самцов крыс разных типологических групп в ходе острого ЭБС. p<0,05, p<0,01, p<0,001 - *, **, *** - по сравнению с исходным состоянием спокойного бодрствования.
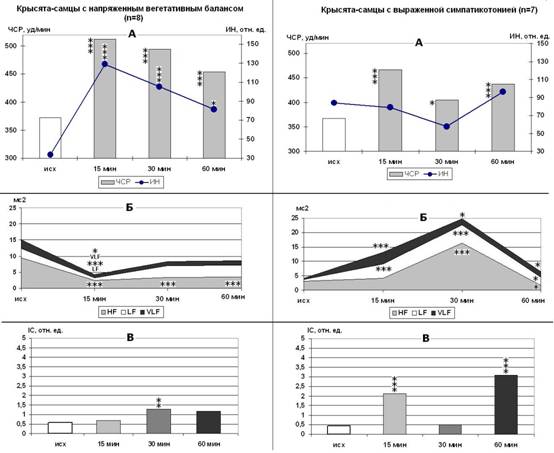
Рисунок 2. Изменения ЧСР и ИН (А), мощности волн спектра ВСР (Б) и IC (В) у крысят-самцов разных типологических групп в ходе острого ЭБС. p<0,05, p<0,01, p<0,001 - *, **, *** - по сравнению с исходным состоянием спокойного бодрствования.
Полученные результаты свидетельствуют, что в работе с нелинейными крысами необходимо придерживаться дифференцированного подхода, учитывающего исходное состояние регуляторных систем, определяемое возрастом, полом и индивидуально-типологическими особенностями организма. Среди самцов крыс «крайними» является симпатотонический тип, а среди самок – ваготонический тип, которые отличаются от других типов по биохимическим показателям. Исходное состояние имеет существенное значение для формирования ответной реакции на острый стресс как со стороны регуляторных систем, так и на уровне тканей в виде изменения интенсивности ПОЛ и антиоксидантной защиты.
3. Влияние периодического введения альфа-токоферола на регуляцию сердечного ритма и показатели свободнорадикальных процессов нелинейных крыс
В связи с обнаружением фактов, указывающих на сопряженность процессов ПОЛ с активностью вегетативных механизмов регуляции, а также в связи с широкой популяризацией витаминно-минеральных комплексов, в том числе, содержащих витамины-антиоксиданты (α-ТФ и др.), представляется необходимым продолжить исследования эффектов длительного приема антиоксидантов на функции организма. Есть данные о том, что α-ТФ проявляет не только антиоксидантные, но и неантиоксидантные свойства (, 2008; и соавт., 1987, 2006; Azzi А., 2007).
Нами установлено, что введение α-ТФ способствует формированию к 42-дневному возрасту более высокой ЧСР у крысят-самцов и очень напряженного СР у крысят-самок (табл. 7). У половозрелых самцов на фоне введения α-ТФ повышена вариабельность СР, что обычно рассматривается как признак роста парасимпатических влияний ( и соавт., 1984, 2001). У самок наблюдается небольшое урежение ЧСР при высокой ригидности СР, что указывает на рассогласование между нервным и гуморальным каналами регуляции. У половозрелых крыс и крысят, получавших α-ТФ, ЧСР коррелирует как с Мо (r= -0,99, р<0,01), так и с показателями нервных влияний: RMSSD (r= -045 и r= -0,68, р<0,01), HF (r= -0,63, р<0,01).
Таблица 7
Показатели ВСР и свободнорадикального гомеостаза нелинейных крыс разного пола и возраста, получавших α-ТФ (M±m)
Показатели ВСР и СРО | 42-дневные | Половозрелые | ||
самцы n=31 | самки n=24 | самцы n=79 | самки n=54 | |
ЧСР, уд/мин | 409,5±4,9 ххх | 414,3±10,7 | 320,4±3,2 ^^^ | 335,6±4,8 х, ^^^, □□ |
Мо, мс | 147,2±1,8 ххх | 148,7±4,2 | 189,5±1,9 ^^^ | 179,5±2,6 х, ^^^, □□ |
RMSSD, мс | 2,9±0,17 х | 2,4±0,25 | 5,7±0,22 ^ | 4,2±0,23 ^^, □□□ |
SD, мс | 4,2±0,43 | 3,1±0,44 | 7,5±0,33 хх, ^^^ | 5,8±0,28 ^^^, □□□ |
ИН, отн. ед. | 82,4±7,2 | 142,1±18,6 хх □□ | 24,5±3,3 ^^^ | 52,9±6,3 х, ^^^, □□□ |
TP, мс2 | 8,11±2,33 | 3,33±0,77 | 21,91±2,85 ^ | 16,86±2,77 ^^ |
HF, % | 56,2±3,48 | 65,2±4,11 х | 47,2±2,13 | 38,7±3,32 □ |
LF, % | 21,2±1,64 | 14,9±2,15 □, х | 29,3±1,39 | 26,3±2,06 |
VLF, % | 22,6±2,27 | 19,8±2,79 | 23,3±3,51 | 34,9±2,94 □ |
IC, отн. ед. | 1,19±0,22 | 0,94±0,19 | 1,20±0,16 | 1,74±0,23 □ |
ТБК-РП пл. | 1,91±0,11 | 2,42±0,25 | 1,74±0,16 ххх | 1,87±0,19 |
Катал. пл. | 22,2±3,8 | 24,6±2,8 | 56,5±5,8 ^^^ | 50,4±4,9 ^^^ |
Катал. эр. | 202,3±12,8 | 184,6±12,3 | 223,6±5,9 | 216,7±8,6 ^ |
АОА пл., % | 70,2±2,4 | 62,3±3,1 х | 81,5±4,1 | 85,1±3,3 ^^^ |
ТБК-РП миок. | 0,66±0,03 | 0,73±0,04 | 0,32±0,04ххх, ^^^ | 0,21±0,03 ххх,^^^, □ |
АЗ-ПОЛ миок. | 8,12±0,66 | 6,48±0,42 □ | 3,21±0,30 ххх, ^^^ | 2,75±0,38 хх, ^ |
Катал миок. | 7,5±0,64 ххх | 5,5±0,90 хх | 8,3±1,2 ххх | 10,9±1,2 ^^ |
АОА миок., % | 68,6±3,2 | 76,7±2,9 | 84,4±2,7 ^^^, х | 88,7±3,8 х, ^ |
ТБК-РП печ. | 0,36±0,05 ххх | 0,30±0,02 ххх | 0,36±0,03 ххх | 0,29±0,01 х |
АЗ-ПОЛ печ. | 4,58±0,99 ххх | 2,42±0,40 ххх | 5,00±0,73 ххх | 2,15±0,14 ххх, □□□ |
Катал печ. | 258,6±2,4 ххх | 251,6±3,9 ххх | 219,6±7,1 ^^^,ххх | 208,9±8,9 ххх, ^^^ |
Примечание. p<0.05, p<0.01, p<0.001 - х, хх, ххх - по сравнению с группами контрольных крыс соответствующего пола и возраста; □, □□, □□□ между самками и самцами каждой возрастной группы, ^, ^^, ^^^ - по сравнению с 42-дневными крысятами соответствующего пола.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
