

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
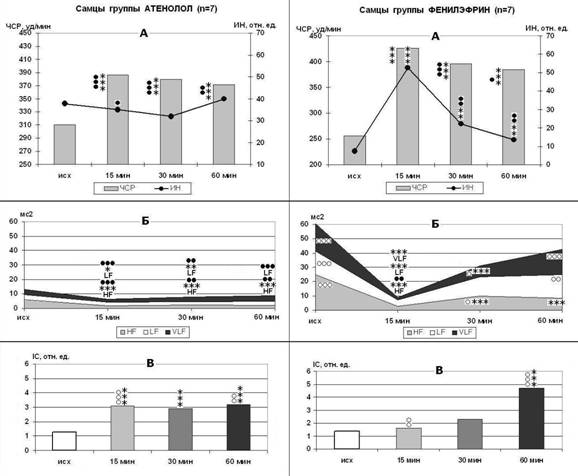
Рисунок 4. Стресс-индуцированные изменения ЧСР и ИН (А), мощности волн спектра ВСР (Б) и IC (В) у самцов крыс в группах Атенолол и Фенилэфрин. p<0,05, p<0,01, p<0,001 - *, **, *** - по сравнению с исходным состоянием спокойного бодрствования; ○,○○,○○○ – выше контроля; ●, ●●, ●●● – ниже контроля.
в исходно недоминирующих областях, что указывает на реализацию эффектов парасимпатической характера, а у крысят и половозрелых самок с ДLF% и ДVLF% - снижается мощность всех волн СР, особенно в доминирующих диапазонах, .то есть проявляются эффекты адренергической направленности.
Эффекты стимуляции α1-АР в отношении ВСР проявляют большую зависимость от возраста и половой принадлежности особей, нежели эффекты блокады β1-АР. Разнонаправленность эффектов стимуляции α1-АР может быть связана с возрастными особенностями их экспрессии и встраивания в мембраны кардиомиоцитов, функционирования систем внутриклеточных посредников ( и соавт., 2011).
Среди крыс с ДHF% блокада β1-АР снижает ЧСР и повышает вариабельность СР только у особей с типом С, что подтверждает высокую выраженность у них симпатического тонуса. На введение ФЭ наибольшее урежение ЧСР и усиление волн ВСР происходит у половозрелых крыс с типами В и ВБ, что связано с высокой активностью парасимпатического канала регуляции у таких крыс. Следовательно, исходный тип регуляции СР определяет выраженность и направленность изменений ЧСР и волновых параметров СР на введение АТ и ФЭ.
Блокада β1-АР снижает уровень продуктов ПОЛ в миокарде и печени самцов крыс (в 3,6-4,8 раза, р<0,001) и крысят (р<0,01 - р<0,001), активность каталазы в тканях (р<0,001) и в плазме крови (в 2-4 раза, р<0,001). Введение агониста α1-АР, напротив, повышает уровень ТБК-РП и каталазную активность в плазме крови половозрелых крыс (в 2,5 раза, р<0,001, на 33,8-42,3%, р<0,05) и крысят (р<0,05, р<0,001).
Блокада β1-АР ограничивает основные проявления стресс-реакции (рис. 4). Стресс-индуцированные изменения СР развиваются по слабо прессорному типу при снижении мощности HF-волн (р<0,001), видимо, за счет торможения активности парасимпатических механизмов со стороны эрготропных зон гипоталамуса и ствола мозга. При этом в плазме крови у самцов и самок увеличивается концентрация ТБК-РП на 50% (р<0,05), активность каталазы в 3,1-4 раза (р<0,001), хотя абсолютные значения этих показателей ниже контрольных (р<0,001).
На фоне введения агониста α1-АР стресс вызывает резкое повышение ЧСР (на 66%-45%, р<0,001) и ИН (в 2-4,3 раза, р<0,001), то есть реакция на стресс носит выраженный взрывной характер (рис. 4). После резкого падения вариабельности СР в начале ЭБС мощность волн LF и VLF нарастает: у самцов при одновременном снижении HF-волн (р<0,001), у самок – одновременно с повышением HF-волн (р<0,001). Однако, изменения параметров ПОЛ выражены умеренно, каталазная активность плазмы увеличивается только на 45-80% (р<0,001). У крысят группы ФЭ стресс вызывает резкое падение мощности волн всех диапазонов спектра (р<0,001 - р<0,05), интенсификацию ПОЛ в миокарде (р<0,01) и рост каталазной активности плазмы в 3,4 раза (р<0,001).
Таким образом, адренергические механизмы через периферические β1- и α1-АР участвуют в формирование амплитуды волн всех частотных диапазонов спектра ВСР, в поддержании фонового уровня ПОЛ и регуляции (или модуляции) активности каталазы в организме крыс разного возраста. Периферические β1-АР являются основным каналом, через который в стрессовых ситуациях стимулируется рост ЧСР и ИН, усиление волн LF и VLF, интенсификация ПОЛ и активация АО-защиты крови и тканей. Блокада β1-АР ограничивает стрессорную мобилизацию функциональных и пластических резервов сердечно-сосудистой системы, делает более жесткой регуляцию СР. Связывание КА с α1-АР в ситуации стресса у половозрелых крыс важно для подержания активности депрессорных механизмов, а у крысят – для усиления адренергической реакции.
Учитывая выше изложенные данные об эффектах α-ТФ, а также результаты других авторов (Liang C., 2000; Manzella D. et al., 2001 и др.), изучали особенности изменений ВСР и ПОЛ при блокаде периферических β1-АР и стимуляции α1-АР у крыс разного пола и возраста, получавших α-ТФ.
Оказалось, что на фоне α-ТФ блокада β1-АР снижает ЧСР (на 6-24%, р<0,05 - р<0,001), повышает вариабельность СР во всех спектральных диапазонах, особенно в исходно недоминирующих, у самцов крыс и крысят (р<0,05 - р<0,001). У самок, напротив, мощность волн СР снижается (р<0,001). На фоне α-ТФ введение агониста α1-АР слабее влияет на ЧСР (р<0,05), но сильнее повышает мощность волн СР у самцов (особенно HF, на 106-232%, р<0,01), а также у самок и крысят (с типом ДHF%) (р<0,001).
Эффекты АТ и ФЭ в отношении ПОЛ и каталазной активности крови и тканей проявляются и на фоне приема α-ТФ, что обусловлено специфическими механизмами их действия. Вместе с тем, на фоне α-ТФ слабее проявились как угнетение каталазной активности крови и тканей при блокаде β1-АР, так и рост ПОЛ и каталазной активности в плазме крови при стимуляции α1-АР. Потенцирующий эффект сочетанного введения α-ТФ и ФЭ проявился только в отношении каталазной активности печени половозрелых крыс (р<0,001).
В условиях стресса у половозрелых крыс, получавших α-ТФ, формируется очень ригидный СР при слабой интенсификации ПОЛ и при блокаде β1-АР, и при стимуляции α1-АР, хотя в первом случае ригидность СР сочетается со слабой тахикардией (у самцов до 14,5%, р<0,001), а во втором – с сильной тахикардией (до 57-47%, р<0,001). У крысят стрессорные изменения выражены сильнее в виде роста мощности HF-волн (р<0,001, в серии α-ТФ+АТ) и VLF-волн (в серии α-ТФ+ФЭ), повышения уровня ТБК-РП и скорости АЗ-ПОЛ в печени (до 30%, р<0,05), роста каталазной активности плазмы (в 4 и 2,4 раза, р<0,001).
Сильный и однонаправленный рост мощности всех волн спектра ВСР у крыс при блокаде β1-АР и стимуляции α1-АР на фоне α-ТФ свидетельствует преобладании механизмов парасимпатической направленности, что согласуется с выше приведенными результатами, а также данными других авторов о повышении парасимпатических влияний на сердце при длительном приеме α-ТФ (Manzella D. et al., 2001; Olatunji L. A., Soladoye A. O., 2008) и изменении проницаемости мембран кардиомиоцитов для ионов К+, Na+ на фоне α-ТФ (, , 1990). Высокая напряженность СР во время стресса, вероятно, потенцирована модулирующим влиянием α-ТФ на морфофункциональное состояние моноаминергических и пептидергических ядер гипоталамуса (, 2008), на обмен КА в организме (Behrens W. A. et al., 1986), обусловлена чувствительностью вегетативных центров ствола мозга и гипоталамуса к уровню свободных радикалов (Campese V. M. et al., 2004; Cardoso L. M. et al., 2006, 2009; Ye S. et al., 2006).
Результаты исследования указывают на способность α-ТФ модулировать эффекты препаратов, влияющих на периферические АР. То есть, активность вегетативных механизмов регуляции влияет на интенсивность перекисных процессов и мощность АО-защиты, но и введение α-ТФ отражается на механизмах регуляции СР и метаболических процессов. Антиоксидантные свойства α-ТФ сильнее проявляются в условиях, способствующих индукции перекисных процессов и ослаблению АО-защиты (при стрессе, при стимуляции периферических адренорецепторов), нежели в условиях фоновой активности.
5. Половые, возрастные, типологические особенности и стресс-индуцированные изменения ВСР и параметров свободнорадикального гомеостаза нелинейных крыс при воздействии на центральные нейромедиаторные процессы
Следующим этапом работы стало изучение роли центральных адренергических механизмов в формировании волн ВСР и интенсивности процессов ПОЛ и антиоксидантной защиты.
Блокада синтеза КА (БСК) снижает ЧСР у большинства самцов и крысят на 6-10,5% (р<0,05 - р<0,01), а также у самок с ДLF% и ДVLF% (р<0,001) (табл. 9), ослабляет VLF-волны и сокращает их долю в спектре, особенно у половозрелых самок и крысят (до 1 мс2 и 0,5 мс2, р<0,001; ниже 15-20%, р<0,05 - р<0,001), способствует росту мощность HF-волн у самцов (р<0,001). В свою очередь, повышение активности центральных КА-ергических систем с помощью БЗМ не изменяет фоновую ЧСР, но сопровождается повышением ИН (р<0,05), мощности VLF-волн (на 43-207%, р<0,05 - р<0,001). У самцов эти изменения являются основными, у самок - они менее специфичны.
БСК снижает ОМН у самок крыс (р<0,05), что вызвано, вероятно, сильным угнетением синтеза КА в организме самок, так как эстрадиол способен угнетать активность тирозингидроксилазы в клетках мозгового вещества надпочечников (Serova L. I. et al., 2005). БСК снижает интенсивности ПОЛ в крови и тканях у самцов (в 2,5-2,9 раза, р<0,001), но способствует ее росту у самок (р<0,05 - р<0,01) и в плазме крови крысят (р<0,001), снижает каталазную активность тканей, особенно миокарда (в 3,1-6,6 раза, р<0,001). БЗМ также угнетает ПОЛ в тканях крыс, особенно в печени (в 4,8-2,9 раза, р<0,001), способствует росту каталазной активности в печени (р<0,05 - р<0,001). БСК и БЗМ оказывают разнонаправленное влияние на каталазную активность плазмы крови: БСК сопровождается ее ростом, особенно у самцов (р<0,05), БЗМ – снижением (на 28%, р<0,05).
БСК ограничивает стрессорный рост ЧСР до 25-13% (р<0,001). Снижение активности центрального звена стресс-системы проявляется в ослаблении (в 3,5 раза у самцов, р<0,01) или поддержании на очень низком уровне у самок и крысят (в 6 и 3,3 раза ниже контроля, р<0,01- р<0,001) VLF-колебаний СРпри усилении и резком доминировании HF-волн в спектре ВСР (р<0,001) (рис. 5). Концентрация продуктов ПОЛ в плазме крови и тканях, скорость АЗ-ПОЛ в тканях (р<0,05 - р<0,001), каталазная активность плазмы крови (в 1,7-2 раза, р<0,001) повышаются, хотя в абсолютном выражении были ниже контрольных (р<0,01). Только у крысят с БСК стресс вызывает резкое повышение каталазной активности плазмы крови (в 4 раза, р<0,001), что могла стать результатом максимальной мобилизации запасов КА из надпочечников.
У крыс с БЗМ стресс вызывает стойкую тахикардию (на 23,8-28,5%, р<0,001), резкий подъем мощности VLF-и LF-волн (в 8,2 и 6,4 раза, р<0,01) у самцов (рис. 5) и выраженную прессорную реакцию с медленным усилением VLF-волн (р<0,01) у самок. Изменения параметров ПОЛ крови и тканей выражены слабо и ниже контрольных (р<0,05 - р<0,001), но рост каталазной активности плазмы сильнее, чем у крыс с БСК: в 2,2-2,9 раза (р<0,001). То есть, при БСК реакция на стресс умеренно ослаблена, а при БЗМ носит взрывной или умеренно прессорный характер.
На основе полученных данных можно заключить, что активность центральных КА-ергических нейронных структур непосредственно проявляется в формировании VLF-волн спектра ВСР и повышение централизации управления СР. Результаты подтверждают гипотезу, согласно которой VLF-волны отражают активность надсегментарных эрготропных структур центрального контура регуляции ( и соавт., 2001; , 2003).
Таблица 9
Изменения параметров ВСР половозрелых и неполовозрелых нелинейных крыс различных типологических
групп при БСК и БЗМ
Показатели ВСР | Типы Груп-пы | Самцы половозрелые | Самки половозрелые | 6-недельные крысята-самцы | ||||||
ДHF% n=25 | ДLF% n=7 | ДVLF% n=9 | ДHF% n=24 | ДLF% n=6 | ДVLF% n=11 | ДHF% n=12 | ДLF% n=4 | ДVLF% n=3 | ||
ЧСР, уд/мин | Исх | 323,7±5,7 | 312,8±5,3 | 324,7±6,2 | 349,0±5,5 | 384,3±6,3 | 336,5±5,4 | 405,4±5,5 | 416,0±6,0 | 409,6±5,4 |
БСК | ↓6,5% □ | - | ↓10,3% □□ | ↓3,2% | ↓16% □□□ | ↓4,3% □□ | ↓9,4% □□ | ↓6,3% □□ | ↓2,6% | |
БЗМ | ↑4,6% □ | - | - | ↓4,3% | ↓7,7% | ↓6,4% □ | нет | нет | нет | |
SD, мс | Исх | 6,3±0,58 | 8,7±0,60 | 8,7±0,62 | 5,5±0,54 | 6,4±0,52 | 6,3±0,28 | 6,6±0,51 | 5,8±0,37 | 7,0±0,58 |
БСК | ↑23,5% | ↓29,4% | ↓20,2% | ↓29,4% □ | ↓40% □□ | ↓38,5%□□□ | ↓48,5%□□□ | ↓65,5%□□□ | ↓81,4%□□□ | |
БЗМ | ↑36,8% □ | ↑25% □ | ↑15,3% | ↑25,8% | ↑10,4% | ↑34,4% □ | нет | нет | нет | |
ИН, отн. ед. | Исх | 24,8±2,87 | 16,2±2,27 | 15,7±1,64 | 33,2±3,83 | 33,6±3,33 | 28,8±2,98 | 48,7±4,10 | 46,1±5,26 | 48,9±4,06 |
БСК | ↓41% □ | ↑104% □ | ↑23,5% | ↑66,2%□□□ | ↑53,8% □ | ↑91,8%□□□ | ↑48% □□ | ↑161%□□□ | ↑465%□□□ | |
БЗМ | ↑30% □ | ↑14% | ↑12% | ↓29,7% □ | ↓29,3% | ↓37,3% □ | нет | нет | нет | |
HF, мс2 | Исх | 9,10±0,70 | 6,20±0,58 | 5,97±0,48 | 6,74±0,63 | 5,18±0,53 | 3,36±0,39 | 5,84±0,53 | 2,93±0,18 | 3,94±0,64 |
БСК | ↑113%□□□ | ↑90% □ | ↑104% □□□ | ↓35,8% □ | ↓10,5% | ↑4,4% | ↓13% | ↓44,8%□□ | ↓78% □□ | |
БЗМ | ↓30% | - | ↓23,7% | ↑135% □□ | ↑78,8% □ | ↑160%□□□ | нет | нет | нет | |
LF, мс2 | Исх | 4,15±0,54 | 11,17±1,20 | 7,39±0,68 | 3,16±0,33 | 9,04±0,66 | 2,28±0,23 | 3,73±0,44 | 6,70±0,47 | 3,84±0,42 |
БСК | ↑22,5% | ↓20,8% | ↓40,5% | ↓28% | ↓40,8% □ | ↑17,7% | ↑8% | ↓96% □□□ | ↓98%□□□ | |
БЗМ | ↑41,4% □ | - | ↑57,8% | ↑109%□□□ | ↓29,8% | ↑85,2%□□ | нет | нет | нет | |
VLF, мс2 | Исх | 3,47±0,47 | 5,64±0,55 | 10,63±0,92 | 2,78±0,40 | 4,17±0,41 | 5,73±0,55 | 1,09±0,10 | 3,58±0,35 | 5,69±0,63 |
БСК | - | ↓40,2% □ | ↓61% □□□ | ↓55,7% □□ | ↓53,9% □□ | ↓73% □□□ | ↓48% □□ | ↓93% □□□ | ↓99% □□□ | |
БЗМ | ↑74% □□ | ↑207% □□ | ↑43% □ | ↑92,3%□□□ | ↑31,7% | ↑88% □□ | нет | нет | нет | |
IC, отн. ед. | Исх | 0,84±0,08 | 2,73±0,36 | 3,02±0,35 | 0,88±0,09 | 2,37±0,18 | 2,61±0,22 | 0,83±0,07 | 3,50±0,27 | 3,09±0,16 |
БСК | ↓48,6% □ | ↓74% □□□ | ↓76% □□□ | ↓17% | ↓38,3% □□ | ↓53% □□□ | ↑9,6% | ↓91% □□□ | ↓95% □□□ | |
БЗМ | ↑111% □□□ | ↑66% □ | ↑92% □□ | ↑1% | ↓48% □ | ↓27,6% □ | нет | нет | нет | |
Формула состоя-ния | Исх | ДHF+СМВ | ДLF+СМВ | ДVLF+СМВ | ДHF+СМВ | ДLF+СМВ | ДVLF+СМВ | ДHF+СМВ | ДLF+СМВ | ДVLF+СМВ |
БСК | ДHF+ВМВ | ДHF+СМВ | ДHF+СМВ | ДНF+НМВ | ДНF+НМВ | ДHF+СМВ | ДНF+СМВ | ДHF+НМВ | ДНF+ОHМВ | |
БЗМ | ДVLF+НМВ | ДVLF+ВМВ | ДVLF+ВМВ | ДHF+ВМВ | ДHF+ВМВ | ДVLF+ВМВ | нет | нет | нет |
Примечание. p<0,05, p<0,01, p<0,001 - □, □□, □□□ – по сравнению с исходным состоянием, значками ↑ и ↓ показано повышение или снижение показателей относительно исходных значений. В группе БСК: ♂ - ДHF 13, ДLF 3, ДVLF 5; ♀- ДHF 12, ДLF 3, ДVLF 6; крысята - ДHF 12, ДLF 4, ДVLF 3; в группе БЗМ: ♂ - ДHF 12, ДLF 4, ДVLF 4; ♀- ДHF 12, ДLF 3, ДVLF 5.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
