

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
В раннем онтогенезе введение α-ТФ снижает интенсивность ПОЛ и способствует росту каталазной активности только в печени (р<0,001) (табл. 7). С наступлением половой зрелости у крыс на фоне α-ТФ низкий уровень ТБК-РП обнаруживается также и в миокарде (р<0,001) и в плазме крови самцов (р<0,001), повышается АОА миокарда (р<0,05), снижается скорость АЗ-ПОЛ (р<0,001). Вместе с тем, каталазная активность в тканях снижается (р<0,001), что может быть следствием общего снижения интенсивности ПОЛ. На этом фоне слабее выражены возрастные и половых различия ПОЛ и АО-защиты.
Среди половозрелых крыс, получавших α-ТФ, на 12-10% снижена доля особей с автономным типом регуляции СР и повышена доля крыс с высокой централизацией управления СР. Причем среди самцов увеличена доля особей с ДLF% (до 24%), что в 2,5 раза выше контрольного уровня, а среди самок – доля особей с ДVLF% (до 33%). Среди крысят, напротив, особей с типом ДHF% больше, чем в контроле (до 74%). На фоне α-ТФ среди крыс с ДHF% на 8-11% больше особей с ваготонией (и самцов, и самок), а среди крыс с ДLF% и ДVLF% на 11-18% больше особей с симпатикотонией. Среди крысят преобладание особей с ВС является более резким, чем в контроле. Особенно высока их доля среди крысят-самок (77%), что выше контрольного на 17%.
В группе α-ТФ самцы-ваготоники имеют самую низкую, а самцы с ВБ - самую высокую ОМН (р<0,001). У самцов-симпатотоников активность ХЭ ниже, чем у крыс других типов (р<0,05). Самцы с ВБ и В, как и в контроле, характеризуются более высокими, а симпатотоники - более низкими значениями параметров ПОЛ в плазме крови и в тканях, но эта закономерность выражена слабее.
Главные особенности реакции на стресс при приеме α-ТФ у половозрелых крыс – сильная и стойкая тахикардия, быстрое и сильное повышение мощности LF и VLF-волн, а у животных-симпатотоников – еще и HF-волн. То есть реакция на острый стресс, особенно у самцов, развивается при значительном усилении влияний со стороны надсегментарных нервных структур. Наряду с этим, стрессорные изменения интенсивности ПОЛ в основном нивелированы, а рост каталазной активности плазмы крови ослаблен почти вдвое от контрольного (р<0,001). Следовательно, причиной стрессогенного роста каталазной активности плазмы является повреждение мембран форменных элементов крови. Стресс-индуцированный рост ЧСР у самцов группы α-ТФ, как и в контроле, сопряжен с падением Мо (r= -0,97, р<0,01), повышением SD (r= 0,44, р<0,05), мощности LF (r= 0,66, р<0,01), каталазной активности плазмы крови (r= 0,62, р<0,01).
В зависимости от исходного состояния у крыс группы α-ТФ развиваются следующие варианты реакции на стресс (рис. 3): 1) взрывной с резким повышением централизации
управления и 2,2-1,7-кратным ростом каталазной активности плазмы (р<0,001) (у крыс с ВБ), 2) сбалансированный с умеренным повышением централизации управления и 2,7-2,8-кратным ростом каталазной активности плазмы (р<0,001) (у крыс с типом С), 3) замедленный с постепенным нарастанием централизации управления и усилением прессорных механизмов, 2,4-2,6-кратным повышением каталазной активности плазмы (р<0,001) (у крыс с типом В). У крысят группы α-ТФ в условиях ЭБС быстро формируется очень ригидный СР, каталазная активность крови повышается в 3,5 - 4,4 раза (р<0,001), ОМН - на 7-21% (р<0,05), что в совокупности указывает на резкое повышение активности адренергических механизмов.
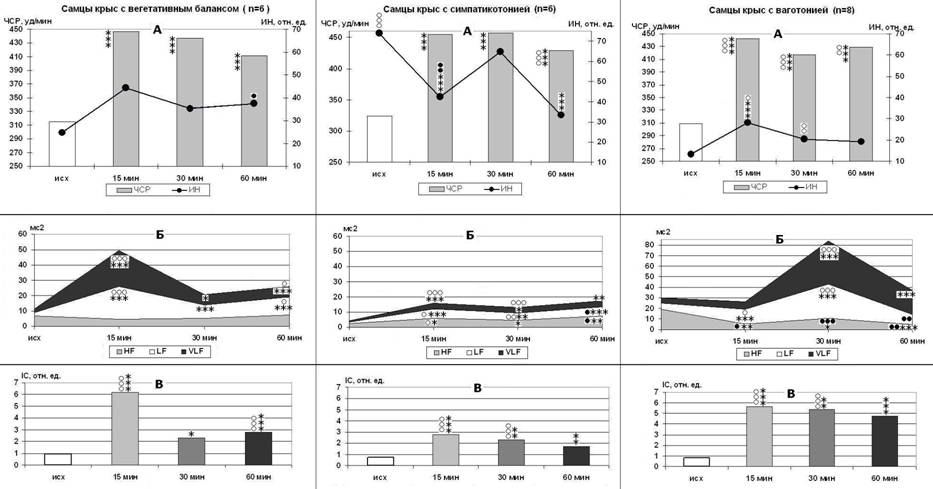
Рисунок 3. Стресс-индуцированные изменения ЧСР и ИН (А), мощности волн спектра ВСР (Б) и IC (В) у самцов крыс с разными типами регуляции в группе α-ТФ. p<0,05, p<0,01, p<0,001 - *, **, *** - по сравнению с исходным состоянием спокойного бодрствования; ○,○○,○○○ – выше соответствующего контроля; ●, ●●, ●●● – ниже соответствующего контроля.
Итак, на фоне α-ТФ существенно изменены взаимоотношения надсегментарного и сегментарного уровней регуляции СР с преобладанием надсегментарных влияний, а у крысят сильно активируются прессорные механизмы регуляции. Повышение фоновой вариабельности СР у крыс с ДHF% свидетельствует о высокой активности парасимпатического канала регуляции при введении α-ТФ, что согласуется с результатами клинических исследований (Manzella D. et al., 2001; Olatunji L. A., Soladoye A. O., 2008).
С учетом известных данных литературы ( и соавт., 2001; , 2003; и соавт., 2003), представленные результаты дают основание говорить о модулирующем влиянии α-ТФ на активность стволовых и гипоталамических структур, периферических отделов ВНС. Модулирующие эффекты α-ТФ на регуляторные механизмы способны реализоваться на различных уровнях организации через изменение: 1) функциональной активности моноаминергических и нейроэндокринных ядер гипоталамуса (, 2008); 2) функционального состояния нейронов в симпатических нервных узлах (, 2003) и обмена КА в организме (Behrens W. A. et al., 1986); 3) чувствительности кардиомиоцитов к вегетативным медиаторам (Bakalova R. et al., 2000; Liang C. et al., 2000; Sayar K. et al., 2000); 4) изменение физико-химических свойств клеточных мембран, что влияет на ионную проводимость мембран кардиомиоцитов ( К, , 1990).
Особенности стресс-индуцированных изменений показателей ВСР и свободнорадикального гомеостаза свидетельствуют о том, что введение α-ТФ потенцирует у половозрелых животных (непосредственно или опосредованно) механизмы, активно противостоящие стрессорным изменениям как на уровне тканевого метаболизма и перекисных процессов, так и на уровне регуляторных систем.
4. Половые, возрастные, типологические особенности и стресс-индуцированные изменения ВСР и параметров перекисного окисления липидов у нелинейных крыс при блокаде и стимуляции периферических адренорецепторов
Вопрос о роли адренергических механизмов регуляции в возникновении волн СР и поддержании фоновой интенсивности СРО до сих пор остается дискуссионным. В этой связи нами исследовано влияние блокады периферических β1-АР атенололом и стимуляции периферических α1-АР фенилэфрином на ВСР, ПОЛ и каталазную активность крови и тканей крыс разного пола и возраста, с учетом типа регуляции СР, определенного у крыс в исходном состоянии.
Систематическое введение блокатора β1-АР урежает ЧСР у самок крыс и крысят (на 4,9 - 11,9%, р<0,05 - р<0,001), имеющих более высокую фоновую ЧСР, чем самцы. Мощность волн СР снижается у всех крыс, наиболее сильно - в исходно доминирующей области спектра (табл. 8). Ослабление доминирующих волн может сочетаться с ослаблением или усилением колебаний на других частотах. Эта закономерность прослеживается отчетливее у самцов, чем у самок. Стимуляция α1-АР сопровождается урежением ЧСР у животных обоих полов и возрастов (на 7,5-18,2%) (табл. 8). При этом у самцов, а также самок с ДHF% повышается вариабельность СР во всех диапазонах спектра, особенно
Таблица 8
Изменения параметров ВСР половозрелых и неполовозрелых нелинейных крыс различных типологических групп при блокаде и стимуляции периферических адренорецепторов
Показатели ВСР | Типы Группы | Самцы половозрелые | Самки половозрелые | 6-недельные крысята-самцы | ||||||
ДHF% n=48 | ДLF% n=8 | ДVLF% n=7 | ДHF% n=42 | ДLF% n=7 | ДVLF% n=10 | ДHF% n=38 | ДLF% n=7 | ДVLF% n=6 | ||
ЧСР, уд/мин | Исх | 318,7±7,0 | 306,6±2,9 | 316,2±5,5 | 345,8±3,3 | 333,9±4,2 | 357,9±4,4 | 410,9±7,2 | 443,0±4,9 | 448,3±7,0 |
АТ | ↓3,2% | ↓1,6% | ↓2,4% | ↓11,3%□□□ | ↓1,7% | ↓4,9% □□ | ↓5,3% | ↓11,9% □□ | ↓10,2% □ | |
ФЭ | ↓18,2%□□□ | ↓18,7%□□□ | ↓15,6%□□□ | ↓7,4% □ | ↓7,4% □ | ↓15,2%□□□ | ↓7,7% □□□ | ↓7,5% □□□ | ↓3,9% | |
SD, мс | Исх | 5,8±0,35 | 7,1±0,50 | 9,6±0,36 | 5,3±0,45 | 7,6±0,48 | 6,1±0,26 | 6,3±0,48 | 6,3±0,29 | 4,7±0,14 |
АТ | ↓11,1% | ↓22,2% □ | ↓64,8%□□□ | - | ↓38,2% □□ | ↓6,4% | ↓13% | ↓28,4% □ | ↓12,5% | |
ФЭ | ↑93,5%□□□ | ↑53,9% □ | ↑62,5% □□ | ↑43,8%□□□ | ↓73,7%□□□ | - | ↓12,3% | ↓9% | ↑24,5% | |
ИН, отн. ед. | Исх | 32,8±2,52 | 17,6±2,31 | 17,7±2,41 | 46,2±4,63 | 26,3±3,08 | 27,9±2,54 | 43,6±4,24 | 40,9±4,9 | 58,9±3,60 |
АТ | ↑18,7% | ↑116% □□ | ↑390%□□□ | ↓14,6% | ↑62,3% □□ | - | ↑19,2% | ↑78% □□ | ↑4% | |
ФЭ | ↓72% □□□ | ↓55% □ | ↓60% □□□ | ↓8,9% | ↓308%□□□ | ↓6,6% | ↑24% | ↓11,5% | ↑10,5% | |
HF, мс2 | Исх | 11,24±1,34 | 8,95±1,15 | 7,18±0,73 | 7,11±0,50 | 6,61±0,58 | 2,61±0,23 | 5,55±0,37 | 3,66±0,52 | 1,84±0,06 |
АТ | ↓42,5% □□ | ↓9,1% | ↓65% □□□ | ↓54,6%□□□ | ↓57% □□ | ↑131% □□ | ↓51% □□□ | ↓25% | ↑227% □□ | |
ФЭ | ↑54,6% | ↑124% □ | ↑88% □ | ↓46,8% □ | ↓88% □□□ | ↑40% □□ | ↓57% □□ | ↑28% | ↓7% | |
LF, мс2 | Исх | 3,65±0,51 | 18,27±2,38 | 8,06±0,76 | 2,40±0,28 | 12,77±1,24 | 3,66±0,30 | 2,93±0,22 | 7,75±0,77 | 2,30±0,18 |
АТ | ↑60% □ | ↓78% □□□ | ↓79% □□□ | ↑120%□□□ | ↓57% □□□ | ↑68% □ | ↓19,3% | ↓71% □□□ | ↑109% □ | |
ФЭ | ↑175% □□□ | ↑60,7% | ↑125%□□□ | ↑88% | ↓93,8%□□□ | ↓65,7%□□□ | ↓43% □ | ↓52,5% □ | ↓61% □□ | |
VLF, мс2 | Исх | 2,72±0,31 | 8,41±1,02 | 12,80±1,15 | 2,73±0,41 | 8,72±0,63 | 6,51±0,41 | 2,30±0,27 | 4,85±0,61 | 3,10±0,18 |
АТ | ↑54% □ | ↓64,1% □□ | ↓87% □□□ | ↑66% □ | ↓34,8% □ | ↓53% □□□ | ↑36% □ | ↓29% □ | ↑107% □□ | |
ФЭ | ↑353% □□□ | ↑108% □ | - | ↑49% □ | ↓93% □□□ | ↓83,6%□□□ | ↑25,4% | ↓53% □□□ | ↓59% □□□ | |
IC, отн. ед. | Исх | 0,56±0,06 | 3,13±0,29 | 3,03±0,31 | 0,78±0,09 | 3,27±0,38 | 3,81±0,46 | 0,98±0,09 | 3,43±0,46 | 2,82±0,20 |
АТ | ↑274% □□□ | ↓26,2% | ↓40,7% □□ | ↑307%□□□ | ↓21,4% | ↓64% □□□ | ↑102%□□□ | ↓45% □□ | ↓38% □□□ | |
ФЭ | ↑120% □□□ | ↓46,4% □ | ↓29% □ | ↑283%□□□ | ↓44,8% □ | ↓82,2%□□□ | ↑54% □ | ↓45% □□□ | ↓55% □□□ | |
Формула состоя-ния | Исх | ДHF+СМВ | ДLF+ВМВ | ДVLF+ВМВ | ДHF+СМВ | ДLF+СМВ | ДVLF+СМВ | ДHF+СМВ | ДLF+СМВ | ДVLF+СМВ |
АТ | ДHF+СМВ | ДHF+ВМВ | ДHF+НМВ | ДLF+СМВ | ДVLF+СМВ | ДHF+СМВ | ДVLF+HМВ | ДHF+СМВ | ДVLF+HМВ | |
ФЭ | ДHF+ВМВ | ДLF+ВМВ | ДLF+CМВ | ДLF+HМВ | ДHF+HМВ | ДHF+HМВ | ДHF+HМВ | ДHF+HМВ | ДHF+HМВ |
Примечание. p<0,05, p<0,01, p<0,001 - □ , □□, □□□ – по сравнению с исходным состоянием, значками ↑ и ↓ показано повышение или снижение показателей относительно исходных значений. В группе АТ: ♂ - ДHF 25, ДLF 5, ДVLF 4; ♀- ДHF 22, ДLF 4, ДVLF 5; крысят - ДHF 20, ДLF 3, ДVLF 3; в группе ФЭ: ♂ - ДHF 23, ДLF 3, ДVLF 3; ♀- ДHF 20, ДLF 3, ДVLF 5; крысят - ДHF 18, ДLF 4, ДVLF 3.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
