

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
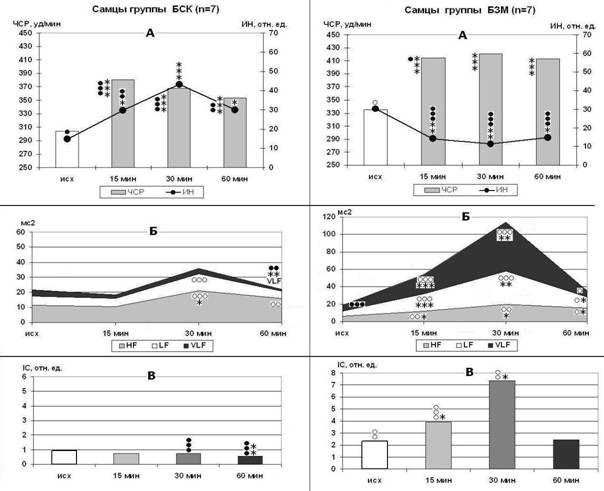
Рисунок 5. Стресс-индуцированные изменения ЧСР и ИН (А), мощности волн спектра ВСР (Б) и IC (В) у самцов крыс с БСК и БЗМ. Обозначения как на рис. 4.
Снижение интенсивности ПОЛ при БСК и БЗМ сильнее выражено у самцов, которые обычно имеют более высокую интенсивность пероксидации. Самки и крысята тяжелее переносят дефицит КА, вероятно, из-за того, что процессы жизнедеятельности протекают у них при более высоком фоновом уровне адренергических влияний. Физиологический уровень КА и активность центральных КА-ергических систем необходимы для обеспечения стресс-индуцированного роста мощности VLF-волн и LF-волн и угнетения дыхательной модуляции СР, для интенсификации перекисных процессов, а также для проявления КА антиоксидантных свойств и модуляции активности компонентов АОС тканей. Результаты исследования показали, что относительная и абсолютная мощность VLF-волн может рассматриваться как показатель активации КА-ергических компонентов центрального звена стресс-системы.
Предварительное введение α-ТФ усиливает эффект БСК в отношении волн VLF (р<0,001). Наряду с этим нормальная ЧСР, высокая напряженность СР, снижение массы надпочечников (р<0,05) и интенсивности ПОЛ в покое (р<0,001), выраженность основных стресс-индуцированных изменений ВСР, ПОЛ и каталазной активности плазмы крови (р<0,05-р<0,001) дают основание полагать, что при формировании БСК на фоне α-ТФ ускоряется развитие сенситизации тканей к КА. Благодаря своим мембранстабилизирующим свойствам (, , 1972; Wang X., Quinn P. J., 1999; и соавт., 2006), α-ТФ способен снизить подвижность АР в мембранах, тем самым потенцировать «временную фиксацию» адренергической реакции. Неуклонное ослабление мощности волн СР в ходе стресса свидетельствует, что дефицит КА делает регуляцию СР менее гибкой, препятствует активации депрессорных механизмов. Следовательно, дефицит КА на фоне α-ТФ способен привести к формированию гиперреактивности прессорных механизмов регуляции за счет роста адреночувствительности периферических органов и тканей.
α-ТФ также потенцирует рост мощности VLF - и LF-волн при БЗМ (на 19-175%, р<0,05-р<0,001), но специфичность эффекта БЗМ в отношении VLF ослаблена из-за повышения общей вариабельности СР и урежения ЧСР. Вместе с тем, формирование взрывной реакции на стресс по параметрам ВСР (р<0,001) и ареактивность к стрессу перекисных процессов в крови и тканях указывают, с одной стороны, на повышение функциональных возможностей регуляторных систем (прежде всего адренергического звена) в отношении СР, а с другой стороны, на развитие десенситизации тканей к КА. Следовательно, модулирующие влияния α-ТФ на эффекты блокады и стимуляции КА-ергических систем могут быть обусловлены ускоренным формированием компенсаторных изменений адренорецепторного аппарата клеток при дефиците или повышенном уровне КА.
ЗАКЛЮЧЕНИЕ
В последние десятилетия неизменно высоким остается интерес к оценке функционального состояния организма, прогнозированию его адаптивных возможностей. В этих целях широко используются методы анализа ВСР (, , 2003; , , 2004; , 2003, 2009; , 2008; , 2009). Поэтому важным является проведение исследований в модельных экспериментах на лабораторных животных для развития представлений о физиологических основах вариабельности кардиоинтервалов, а также для выявления её сопряженности с другими функциями организма, в том числе, с метаболическими процессами.
Анализ ВСР нелинейных крыс выявил, что периоды волн сердечного ритма у этих животных почти в 10 раз короче, чем у человека, поэтому на КИГ крыс длительностью в 5 мин и даже 1 мин хорошо просматриваются все основные периодические составляющие вариабельности СР. Резкое ослабление волн СР под наркозом существенно затрудняет выявление изменений, вызванных экспериментальными воздействиями. В этой связи изучение ВСР с применением спектрального анализа имеет смысл проводить в состоянии бодрствования.
Результаты экспериментов с применением наркоза, с блокадой и стимуляцией центральных моноаминергических структур показали, что частотные характеристики волн СР определяются активностью уровней нервной регуляции. В свою очередь, пробы с изменением положения тела в пространстве, с блокадой и стимуляцией периферических АР свидетельствуют, что амплитуда колебаний кардиоинтервалов на той или иной частоте определяется активностью и взаимодействием симпато-парасимпатических механизмов сегментарного уровня регуляции.
На основе анализа информационной значимости параметров ВСР с применением статистических методов (факторный анализ, дисперсионный анализ, корреляционный анализ) и данных литературы нами разработана «концептуальная модель факторов, определяющих вариабельность СР» (рис. 6). Согласно сути факторного анализа, факторные нагрузки отдельных переменных (показателей ВСР) представляют корреляции между переменными и факторами. ЧСР и Мо кардиоинтервалов (r= -0,97-0,99, p<0,01) образуют самостоятельный фактор F1. Согласно (1978), Мо – наиболее вероятный уровень активности водителя ритма сердца, показатель гуморальных влияний на сердце. Мо слабо сопряжена с другими параметрами ВСР, поэтому считаем, что она отражает основы ритмогенеза сердца – автоматическую активность и электротоническое взаимодействие клеток водителя ритма, которое, согласно (1997), является механизмом синхронизации и формирования устойчивого ритма сердечных сокращений. Учитывая это, мы определили F1 как фактор миогенно-гуморальных механизмов. Влияния вегетативных медиаторов вызывают десинхронизацию между клетками синусно-предсердного узла (, 1997). Это представляется главной причиной вариабельности кардиоинтервалов. Все параметры ВСР, кроме Мо и ЧСР, формируют 2 фактора. Сильные корреляции с F2 имеют показатели ВПМ и абсолютной мощности волн всех частот спектра ВСР. Основываясь на известных физиологических трактовках показателей ВПМ, мы определили F2 как фактор симпато-парасимпатических отношений. F3 образуют нормированные мощности волн СР, в связи с чем он определен как фактор уровней нервной регуляции. Все три фактора представляют собой основные информационные компоненты, заключенные в параметрах ВСР.
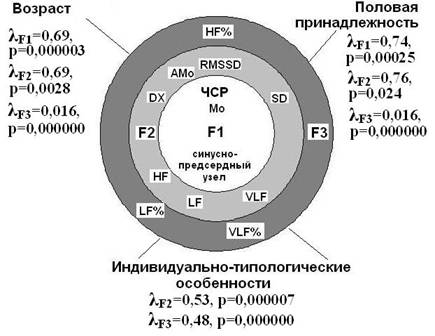
Рисунок 6. Факторы, определяющие вариабельность сердечного ритма.
F1 – фактор миогенно-гуморальных механизмов, F2 – фактор симпато-парасимпатических отношений, F3 – фактор уровней нервной регуляции.
λ - критерий Wilks - результаты дисперсионного анализа (ANOVA/МANOVA results) влияния возраста, пола и индивидуально-типологических особенностей на параметры ВСР.
В соответствии с представленной моделью, наиболее полная характеристика типа регуляции СР может быть дана на основе выявления доминирующего спектрального диапазона (определение ведущего уровня регуляции) и абсолютной мощности колебаний кардиоинтервалов на доминирующей частоте (определение характера симпато-парасимпатических отношений).
Применение единого методологического подхода к регистрации и анализу ВСР у бодрствующих нелинейных крыс позволило нам выявить основные закономерности возрастных изменений волновых характеристик СР, половые и типологические особенности ВСР. То есть на СР оказывают влияние такие факторы как возраст, пол и индивидуально-типологические свойства организма. Дисперсионный анализ (ANOVA/МANOVA, Statistica 6.0) выявил, что на ЧСР и Мо наибольшее влияние оказывают возраст и половая принадлежность, симпато-парасимпатические отношения в набольшей мере зависят от индивидуально-типологических особенностей, затем от возраста и менее – от половой принадлежности. Активность уровней регуляции практически в равной мере и весьма сильно зависит от пола и возраста, а также от индивидуально-типологических особенностей. Это свидетельствует о необходимости учета возраста, половой принадлежности и исходного состояния регуляторных систем при организации экспериментов на нелинейных крысах.
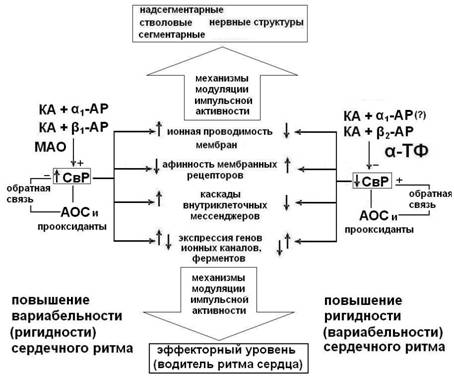
Рисунок 7. Гипотетическая модель взаимосвязей механизмов регуляции сердечного ритма и свободнорадикальных процессов. СвР – свободные радикалы, АОС – антиоксидантная система, МАО – моноаминоксидаза.
В ходе исследований выявлена сопряженность интенсивности процессов свободнорадикального гомеостаза и механизмов вариабельности СР, которая проявилась: а) в повышении общей мощности волн СР и интенсивности ПОЛ у самцов крыс по мере роста и развития организма, б) в более низкой вариабельности СР и низкой интенсивности процессов пероксидации липидов у самок по сравнению с самцами, у особей-симпатотоников по сравнению с другими типами регуляции СР, в) в более высокой интенсивности процессов пероксидации в раннем онтогенезе и ее более значительном снижении с возрастом у крыс с высокими симпатическими влияниями на СР.
Дисперсионный анализ (ANOVA/МANOVA, Statistica 6.0) выявил, что интенсивность ПОЛ и каталазная активность крови и тканей в наибольшей мере зависит от возраста λ=0,010-0,14-0,46 (р<0,000000), от половой принадлежности λ=0,47-0,71-0,66 (р<0,000001-р<0,000005), менее – от типа регуляции СР λ=0,708-0,762 (р<0,003-р<0,03).
В экспериментах с блокадой и стимуляцией периферических и центральных адренергических структур показано участие адренергических механизмов регуляции в поддержании фоновой интенсивности ПОЛ и каталазной активности крови и тканей крыс разного возраста и пола. Выявлены сильные корреляции между стресс-индуцированным ростом ЧСР и повышением каталазной активности плазмы крови, а также уровнем продуктов ПОЛ в печеночной ткани у крыс с интактными адренергическими механизмами, и ослабление этих корреляций при блокаде адренорецепторов и синтеза катехоламинов.
Взаимосвязи регуляции СР и механизмов про - и антиоксидантного баланса реализуются через модулирующее влияние α-ТФ на свободно-радикальный гомеостаз и на активность нервных механизмов регуляции СР.
Гипотетические механизмы модулирующего влияния свободных радикалов и α-ТФ на импульсную активность нейронов и клеток водителя ритма сердца представлены на рис. 7. Модуляция ионной проводимости мембран, сродства клеточных рецепторов к лигандам посредством окислительной модификации белков и липидов способна потенцировать снижение или повышение вариабельности или ригидности сердечного ритма. Неоднозначность модулирующих эффектов определяется функционированием сложной системы антиоксидантов-прооксидантов, которая постоянно регулирует наработку свободных радикалов в нейронах и клетках сердца.
ВЫВОДЫ
1. На основе проведенных исследований сформулированы представления о возрастных, половых и типологических особенностях сердечного ритма нелинейных крыс, получены новые данные о природе VLF-компоненты ВСР, показана роль периферических и центральных звеньев адренергических механизмов в регуляции сердечного ритма и процессов пероксидации, сформированы концептуальные представления о взаимосвязях механизмов регуляции сердечного ритма и свободнорадикальных процессов.
2. Основными тенденциями возрастных изменений хронотропной функции сердца нелинейных крыс в период от рождения до половой зрелости являются урежение ЧСР и повышение мощности волн сердечного ритма, особенно HF. Критическими для становления симпатоадреналовых механизмов и надсегментарных влияний является 21-дневный возраст, а для становления парасимпатических механизмов - 28-дневный возраст, к 35-му дню жизни формируется «взрослый» тип волновой структуры сердечного ритма.
3. Самки крыс имеют более высокую ЧСР, чем самцы, как в 42-дневном возрасте, так и в период половой зрелости, что указывает на преобладание симпатоадреналовых механизмов в регуляции сердечного ритма у особей женского пола. Половозрелые самки отличаются от самцов низкой мощностью HF-волн и более высокой мощностью VLF-волн. Половые различия в регуляции сердечного ритма проявляются в реакциях на острый стресс и введение фармакологических препаратов, то есть являются функциональными. У особей мужского пола взаимодействия структур центрального и автономного контуров регуляции сердечного ритма носят преимущественно реципрокный характер, а у особей женского пола – взаимно компенсационный характер.
4. Среди половозрелых нелинейных крыс, находящихся в состоянии спокойного бодрствования, присутствуют животные 9 типов регуляции СР, которые характеризуются различной активностью уровней нервной регуляции, дифференцируемой по нормированной мощности спектров ВСР (ДHF% - 62%, ДLF% - 14%, ДVLF% - 24%), и различными симпато-парасимпатическими отношениями, оцениваемыми по абсолютным мощностям волн доминирующей области спектра, которые могут быть низкими, средними и высокими. Среди 42-дневных крысят также присутствуют особи с различной активностью уровней нервной регуляции ВСР, но только 2 типов симпато-парасимпатических отношений: с очень низкой (выраженная симпатикотония) и средней (напряженный вегетативный баланс) абсолютной мощностью волн сердечного ритма.
5. Животные с разными типами регуляции сердечного ритма различаются по интенсивности ПОЛ и мощности антиоксидантной защиты крови и тканей: самцы с симпатикотонией имеют наиболее низкий уровень СРО и каталазной активности тканей. Среди самок ваготоники характеризируются низкой интенсивностью ПОЛ в тканях и высокой каталазной активностью плазмы крови и миокарда. У крысят с выраженной симпатикотонией уровень ПОЛ в тканях выше, чем у особей с напряженным вегетативным балансом.
6. Типологический подход к анализу изменений ВСР, ПОЛ и каталазной активности плазмы крови позволил выявить 3 варианта реакции половозрелых крыс на острый стресс: сбалансированный прессорно-депрессорный, взрывной прессорно-депрессорный, замедленный (депрессорно-прессорный у самцов и прессорно-депрессорный у самок), у 6-недельных крысят - 2 варианта реакции: сбалансированный прессорно-депрессорный и взрывной депрессорно-прессорный.
7. На фоне периодического введения α-токоферола наряду с изменением интенсивности пероксидации липидов обнаружено повышение ригидности сердечного ритма у 6-недельных крысят, увеличение доли особей с высокой активностью структур центрального контура регуляции и снижение доли особей с автономным типом регуляции среди половозрелых крыс, нарастание общей вариабельности кардиоинтервалов у самцов, урежение ЧСР у самок. Стресс-реакция на фоне введения α-токоферола у половозрелых крыс протекает с повышением вариабельности кардиоинтервалов в области LF и VLF, особенно у самцов, а у крысят, напротив, с резким ростом ригидности сердечного ритма.
8. Регуляторные влияния через периферические β1- и α1-адренорецепторы прямо или опосредованно участвуют в формировании волн различных диапазонов спектра ВСР, поддержании фонового уровня ПОЛ и каталазной активности в крови и тканях крыс разного возраста. Блокада β1-адренорецепторов ослабляет стресс-индуцированные изменения ЧСР и ВСР, ПОЛ и каталазной активности крови, стимуляция периферических α1-адренорецепторов ускоряет и усиливает развитие стрессорных изменений ЧСР, волновых параметров сердечного ритма, ПОЛ крови, в меньшей степени – тканей у крыс разного возраста.
9. Блокада синтеза катехоламинов снижает, а стимуляция активности центральных моноаминергических систем введением амитриптилина повышает мощность и долю VLF-волн в спектре ВСР. Это свидетельствует о ведущей центральных катехоламинергических систем в формировании VLF-волн сердечного ритма. Угнетающий эффект блокады синтеза катехоламинов в отношении волн VLF сильнее проявляются у самок крыс и крысят, а также в ситуации острого стресса. Блокада синтеза катехоламинов снижает фоновую интенсивность ПОЛ и каталазную активность тканей, в условиях стресса дефицит катехоламинов потенцирует интенсификацию процессов пероксидации липидов.
10. На фоне введения α-токоферола изменяются эффекты блокады и стимуляции адренергических структур на регуляцию сердечного ритма, что может быть следствием его потенцирующего влияния на развитие сенситизации-десенситизации тканей к катехоламинам. На фоне α-ТФ становятся менее специфичными эффекты блокады синтеза катехоламинов и блокады обратного захвата медиаторов в отношении волн VLF, а острый стресс сопровождается формированием ригидного сердечного ритм и при блокаде, и при стимуляции адренергических механизмов регуляции. α-Токоферол оказывает модулирующее влияние на интенсивность ПОЛ и каталазную активность крови и тканей, измененные в условиях адренергической дисрегуляции.
11. Функциональные взаимосвязи механизмов регуляции сердечного ритма и про-и антиоксидантного баланса осуществляются через участие адренергических влияний в поддержании фонового уровня ПОЛ и каталазной активности крови и тканей, обеспечение их изменений в условиях стресса, через модулирующее влияние α-токоферола на свободно-радикальные процессы и активность нервных механизмов регуляции сердечного ритма. Вероятным уровнем реализации функциональных взаимосвязей являются мембраны нервных и миокардиальных клеток, несущие рецепторы к вегетативным медиаторам и гормонам, с которыми сопряжены каскады внутриклеточных посредников и ионные каналы.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
Монография
1) Вегетативная регуляция сердечного ритма: результаты и перспективы исследований (монография). - Астрахань: ИД «Астраханский университет», 2008. – 140 с. – ISBN 0220-3.
Публикации в изданиях, рекомендованных ВАК РФ
2) Влияние α-токоферола и физической тренировки на регуляцию сердечного ритма и вегетативный баланс у крыс с дефицитом симпатических нервных влияний // Известия вузов. Северо-Кавказский регион. Естественные науки. – 2007. - №4. – С.66-69. (Соавт. , )
3) К вопросу о методах анализа сердечного ритма и типах его регуляции на этапах онтогенеза белых крыс // Естественные науки. Журнал фундаментальных и прикладных исследований – 2008. - №4(25). – С.82-89.
4) Особенности свободнорадикальных процессов у нелинейных крыс с различным типом вегетативной регуляции сердечного ритма: онтогенетический аспект // Известия Самарского НЦ РАН. – 2008. - Спец. выпуск. - Т.2. – С. 84-90. (Соавт. , , )
5) Вариабельность сердечного ритма нелинейных крыс при периодическом введении а-токоферола // Бюллетень эксп. биол. и медицины. – 2009. – Т.147, № 2. – С.130-133.
6) Половые и возрастные особенности вариабельности сердечного ритма и перекисного окисления липидов у крыс при десимпатизации и введении α-токоферола // Известия вузов. Северо-Кавказский регион. Естественные науки. – 2009. - №3. – С.85-90. (Соавт. , )
7) Влияние блокады синтеза катехоламинов на регуляцию сердечного ритма и свободнорадикальные процессы у самцов и самок крыс в условиях покоя и острого стресса // Естественные науки. Журнал фундам. и прикладных исследований. – 2009. - №2 (27). – С.123-130. (Соавт. )
8) К вопросу о применении спектральных и статистических параметров вариабельности сердечного ритма для оценки нейровегетативного состояния организма в эксперименте // Бюллетень СО РАМН. – 2009. – Т. 140, № 6. – С. 30-37.
9) Вариабельность сердечного ритма самцов и самок нелинейных крыс и ее изменения при блокаде синтеза катехоламинов // Бюллетень СО РАМН. – 2009. – Т. 140, № 6. – С. 38-42. (Соавт. )
10) Влияние альфа-токоферола на регуляцию сердечного ритма нелинейных крыс: возрастной и половой аспекты // Известия Самарского научного центра РАН. – 2009. – Т.11, №1(5). – С.979-983.
11) Влияние центральных нейромедиаторных процессов на вариабельность сердечного ритма нелинейных крыс в покое и в условиях острого стресса: к вопросу о природе очень медленноволновой компоненты спектра // Бюллетень эксп. биол. и медицины. – 2010. – Т.149, № 1. – С.14-17. (Соавт. )
12) Модулирующее влияние альфа-токоферола на изменения вариабельности сердечного ритма при введении атенолола: роль исходного состояния регуляторных систем // Сибирский медицинский журнал. – 2010. – Том 25, №2, выпуск 1. – С. 188-189.
13) Влияние агониста α1-адренорецепторов на вариабельность сердечного ритма самцов и самок белых крыс // Естественные науки. Журнал фундаментальных и прикладных исследований. – 2010. - №3 (32). – С. 98-106.
14) Влияние α-токоферола на регуляцию сердечного ритма нелинейных крыс в условиях острого стресса // Известия Самарского научного центра РАН. – 2010. – Т.12, №1(8). – С..
15) Влияние наркоза и рефлекторной стимуляции симпатоадреналовой и парасимпатической систем на вариабельность сердечного ритма крыс // Естественные науки. Журнал фундаментальных и прикладных исследований. – 2011. - №2 (35). – С. 140-148.
16) Типологический подход к оценке результатов фармакологических проб при изучении механизмов регуляции сердечного ритма // Естественные и технические науки. – 2011. - №4. – С.208-211.
17) Влияние альфа-токоферола на эффекты блокады и стимуляции адренергических механизмов регуляции в отношении вариабельности сердечного ритма нелинейных крыс // Естественные и технические науки. – 2011. - №4. – С.212-216. (Соавт. )
18) Влияние адренергической дисрегуляции на свободнорадикальный гомеостаз организма в эксперименте // Естественные науки. Журнал фундаментальных и прикладных исследований. – 2011. - №3 (36). – С. 119-128.
Публикации в других изданиях
19) Влияние токоферола, физической тренировки и их сочетания на гистоструктуру симпатических ганглиев и частоту сердечного ритма белых крыс в условиях десимпатизации гуанетидином // Журнал фундаментальных и прикладных исследований «Естественные науки». - №6. – Астрахань, 2003. - С.69-78. (Соавт. , )
20) Особенности показателей сердечного ритма десимпатизированных крыс при введении α-токоферола, физической тренировке и их сочетании // Вариабельность сердечного ритма: Теоретические аспекты и практическое применение. Тез. докладов междунар. симпозиума. – Ижевск: Изд. Удм. Ун-та. – 2003. – С.95-96. (Соавт. , )
21) Возрастные изменения показателей сердечного ритма в условиях формирования дефицита симпатических нервных влияний, введения a-токоферола и их сочетания // XIX съезд Физиологического общества им. / Материалы съезда. 19-24 сентября 2004 г. – Екатеринбург, 2004. – С.142-143. (Соавт. , )
22) Половые особенности регуляции хронотропной функции сердца крыс при формировании дефицита симпатических нервных влияний и физической тренировке // Научные труды I Съезда физиологов СНГ. Сочи, Дагомыс. 19-23 сентября 2005. – Том 2. – Москва: Медицина-Здоровье. – 2005. – С. 70-71. (Соавт. , )
23) Влияние α-токоферола на стресс-реактивные изменения хронотропной функции сердца белых крыс // Успехи современного естествознания. - №12. – 2005. – С.44-45.
24) Активность каталазы крови при эмоционально-болевом стрессе: эффект витамина Е // Материалы Международной научной конференции «Свободные радикалы, антиоксиданты и старение» - Астрахань, 2006. – С.83-85. (Соавт. , , )
25) Особенности регуляции сердечного ритма у белых крыс при введении альфа-токоферола // Материалы Международной научной конференции «Свободные радикалы, антиоксиданты и старение», Астрахань, 2-3 ноября 2006 г. - Астрахань, ИД «Астраханский университет», 2006. – С.150 – 153.
26) Влияние адаптации к физической нагрузке на стресс-реактивные изменения сердечного ритма самцов и самок белых крыс // Эколого-физиологические проблемы адаптации: Материалы XII Междунар. симпозиума. – М.: РУДН, 2007. – С.257-259.
27) Половые особенности перекисного окисления липидов у крыс при эмоционально-болевом стрессе // Перспектива –2007: Материалы Междунар. конгресса студ., асп. и молодых ученых. – Нальчик: Каб-Балк. Ун-т, 2007. – С.34-36. (Соавт. , , )
28) Оценка разнообразия нелинейных белых крыс по параметрам вариабельности сердечного ритма // Материалы международной научно-практической конференции «Экология биосистем: проблемы изучения, индикации и прогнозирования» 20-25 августа 2007 г. – Астрахань, ИД «Астраханский университет», 2007. – Том 2. - С. 202-205. (Соавт. , )
29) Особенности свободнорадикальных процессов у нелинейных крыс с различным типом регуляции сердечного ритма // Фундаментальные исследования в медицине и биологии: Сб. научных трудов. – Вып.3. – Ставрополь, Изд. СевКавГТУ, 2007. – С.40-47. (Соавт. , С, )
30) Типы регуляции сердечного ритма самцов и самок нелинейных белых крыс в условиях покоя и острого стресса // Естественные науки. Журнал прикладных и фундаментальных исследований. – Астрахань: ИД «Астраханский университет», 2007. - №2 (19). – С. 48-56.
31) Половые особенности вариабельности сердечного ритма нелинейных крыс в возрастном аспекте // Тезисы докл. VI Сибирского физиологического съезда. – Барна8. – В 2 томах. Т.1. – С.46. (Соавт. )
32) Спектральный анализ регуляции сердечного ритма на этапах онтогенеза нелинейных белых крыс // Физиологические механизмы адаптации растущего организма. Тезисы IX Всероссийской научно-теоретической конференции, посвященной юбилею профессора . 3-5 октября 2008 г. – Казань, 2008. – 75-76. (Соавт. )
33) Возрастные особенности регуляции сердечного ритма крыс при эмоционально-болевом стрессе // Физиологические механизмы адаптации растущего организма. Тезисы IX Всероссийской научно-теоретической конференции, посвященной юбилею профессора . 3-5 октября 2008 г. – Казань, 2008. – 77-78. (Соавт. )
34) Влияние альфа-токоферола на стресс-реактивные изменения сердечного ритма белых крыс // Физиология и здоровье человека. Тезисы докладов II Съезда физиологов СНГ. 29-31 октября 2008 г. – Молдова, Кишинев. – 2008. – с.118. (Соавт. )
35) Возрастные особенности взаимосвязей между показателями сердечного ритма и интенсивности свободно-радикальных процессов у нелинейных крыс // Механизмы функционирования висцеральных систем. VI Всероссийская конференция с международным участием, посвященная 50-летию открытия мембранного пищеварения. Тезисы докл. – СПб.: Институт физиологии им. РАН. – 2008. – С.109-110.
36) Изменения сердечного ритма белых крыс при различных режимах введения бета-блокатора: роль исходного состояния регуляторных систем // Вариабельность сердечного ритма: Теоретические аспекты и практическое применение. Материалы IV Всеросс. симп. / Отв. ред. , . – Ижевск, УдГУ, 2008. – С.170-173.
37) Спектральные характеристики сердечного ритма наркотизированных крыс при рефлекторной стимуляции симпатоадреналовых и парасимпатических влияний // Вариабельность сердечного ритма: Теоретические аспекты и практическое применение. Материалы IV Всеросс. симп. / Отв. ред. , . – Ижевск, УдГУ, 2008. – С.174-177.
38) Влияние блокады β1-адренорецепторов на стресс-индуцированные изменения регуляции сердечного ритма и интенсивности перекисного окисления липидов у нелинейных крыс // Вестник РГМУ. Периодический медицинский журнал. – М.: ГОУ ВПО РГМУ Росздрава. – 2009. - №3. – С.42-43. (Соавт. , , )
39) Влияние блокады бета1-адренорецепторов на свободнорадикальные процессы и активность каталазы у самцов и самок белых крыс в условиях покоя и острого стресса // Фундаментальные исследования в биологии и медицине: Сб. науч. трудов. – Вып. 6. – Ставрополь: Изд. СевКавГТУ, 2009. – С. 68-80. (Соавт. , , )
40) Влияние антиоксиданта, антидепрессанта и их комбинации на стресс-индуцированные изменения нейровегетативной регуляции сердечного ритма нелинейных крыс // Нейронаука для медицины и психологии: 5-й Междунар. междисципл. конгресс. Судак, Украина, 3-13 июня 2009 г. – М.: МАКС Пресс, 2009. – С. 137-138. (Соавт. )
41) Половые особенности вариабельности сердечного ритма и интенсивности свободнорадикальных процессов у нелинейных крыс при фармакологической блокаде адренергического звена регуляции // Механизмы функционирования висцеральных систем: VII Всероссийская конференция с междунар. участием, С.-Петербург, 29 сентября - 2 октября 2009 г.: Материалы конференции. – С-Пб., Институт физиологии им. РАН, 2009. – С.236-237.
42) Влияние альфа-токоферола на эффекты β1-адреноблокады в отношении хронотропной функции сердца, про - и антиоксидантной активности крови самцов и самок нелинейных крыс // 2-й Научно-практический симп. «Свободнорадикальная медицина и антиоксидантная терапия», Волгоград, 13-15 мая 2009 г. - Кислород и антиоксиданты. – 2009. - №1. – С. 79-80.
43) Типологические особенности стресс-индуцированых изменений регуляции сердечного ритма и параметров свободнорадикального гомеостаза у крыс разного возраста // Биологические исследования. – 2009. - № 4 (4). – С. 27-40.
44) Влияние исходного состояния регуляторных систем на изменения вариабельности сердечного ритма при введении атенолола // Сборник тезисов докладов Междунар. конгресса «Кардиология на перекрестке наук». – Тюмень. – 2010. – С.161-162.
45) Особенности эффектов препаратов, снижающих симпатоадреналовые влияния, на стресс-индуцированные изменения нейровегетативной регуляции сердечного ритма // Нейронаука для медицины и психологии: 6-й Междунар. междисципл. конгресс. – Судак, Украина, 5-15 июня 2010 г. – М.: МАКС Пресс, 2010. – С.185-186. (Соавт. )
46) Влияние альфа-токоферола на сердечный ритм и каталазную активность крови при блокаде синтеза катехоламинов у самцов нелинейных крыс // Кислород и антиоксиданты. – 2010. - №2. – С.15-16.
47) Влияние антагониста и агониста адренорецепторов на вариабельность сердечного ритма крыс // XXI съезд Физиологического общества им. . Тезисы докладов. – Москва – Калуга: Типография -принт», 2010. – С. 336.
48) Изменения вариабельности сердечного ритма при блокаде синтеза катехоламинов: половой и возрастной аспекты // Нейронаука для медицины и психологии: 6-й Междунар. междисципл. конгресс. – Судак, Украина, 3-13 июня 2011 г. – М.: МАКС Пресс, 2011. – С.253-254. (Соавт. )
49) Стресс-индуцированные изменения каталазной активности плазмы крови при фармакологической блокаде и стимуляции адренергических механизмов регуляции // Материалы II Междунар. научной конференции «Свободные радикалы, антиоксиданты и старение», Астрахань, 2-3 ноября 2011 г. - Астрахань, ИД «Астраханский университет», 2011. – С.93 – 97.
50) Типы регуляции сердечного ритма у нелинейных крыс и их особенности при длительном приеме альфа-токоферола // Вариабельность сердечного ритма: Теоретические аспекты и практическое применение // Материалы V всеросс. симп. / Отв. ред. , . – Ижевск, УдГУ, 2011. – С.423-426.
ПРИНЯТЫЕ СОКРАЩЕНИЯ
α-ТФ – альфа-токоферол
АЗ-ПОЛ – аскорбатзависимое ПОЛ
АО – антиоксидант
АОА – антиокислительная активность
АОС – антиоксидантная система
АР – адренорецепторы
АТ – атенолол
БЗМ – блокада захвата медиаторов
БСК – блокада синтеза катехоламинов
В – ваготония
ВБ - - вегетативный баланс
ВМВ высокая мощность волн СР
ВС - - выраженная симпатикотония
ВСР – вариабельность сердечного ритма
ДHF% - доминирование HF-волн
ДLF% - доминирование LF-волн
ДVLF% - доминирование VLF-волн
КА – катехоламины
НВБ – напряженный вегетативный баланс
НМВ – низкая мощность волн СР
ОМН – относительная масса надпочечников
ПОЛ – перекисное окисление липидов
С – симпатикотония
СМВ – средняя мощность волн СР
СР – сердечный ритм
ТБК-РП – ТБК-реактивные продукты
ФЭ – фенилэфрин
ХЭ – холинэстераза
ЭБС – эмоционально-болевой стресс

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
