

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
- 1 группа образуется видами-эдификаторами. Растения, входящие в данную группу, имеют схожие с Angelica sylvestris требования к условиям экотопа – это крупнотравный вид Filipendula ulmaria, образующая плотные кочки Deschampsia caespitosa, Alopecurus pratensis и Anthriscus sylvestris. Наши исследования показали, что с увеличением проективного покрытия Angelica sylvestris их участие в напочвенном покрове равномерно снижается (рис 1). Минимальные значения покрытия эти виды демонстрируют при максимальных значениях проективного покрытия Angelica sylvestris, т. е. между этими видами и дудником существует отрицательная связь. Она может быть объяснена ярко выраженной конкуренцией между Angelica sylvestris и Filipendula ulmaria, Deschampsia caespitosa, Alopecurus pratensis, Anthriscus sylvestris и способностью дудника подавлять их развитие. В работе Винокуровой (Винокурова,1991) было показано наличие активного эфирного масла в подземных и наземных органах Angelica sylvestris, содержащего терпеноиды, которые накапливаются в почве и, возможно, оказывают ингибирующее аллелопатическое воздействие на окружающие виды- эдификаторы, проявляющееся в уменьшение их проективного покрытия.
Рисунок 2. Изменение проективного покрытия видов эдификаторов при увеличении покрытия Angelica sylvestris
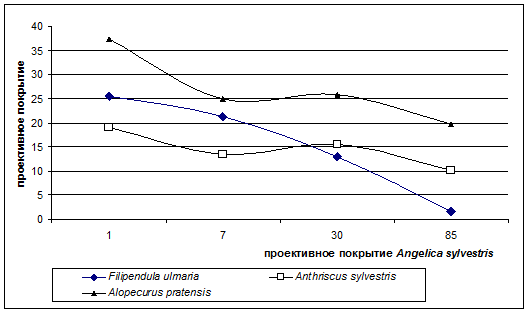
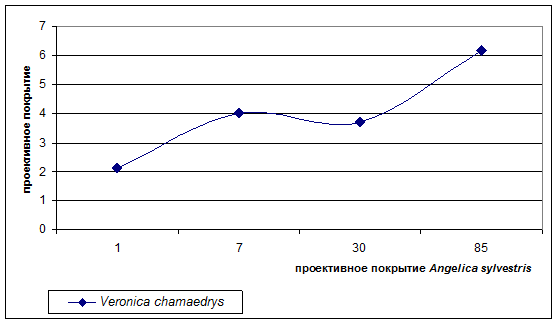
- 2 группа образуется из луговых видов, которые в сообществах скорее являются патиентами. Это такие виды как Poa nemoralis, Veronica chamaedrys, Carex nigra, Geum rivale. График распределения проективного покрытия Veronica chamaedrys (рис.3), показывает, что в напочвенном покрове участие этого вида возрастает с увеличением проективного покрытия Angelica sylvestris. Poa nemoralis, Veronica chamaedrys обнаруживают самые высокие значения проективного покрытия на площадках с проективным покрытием Angelica sylvestris 60-90%. Такой результат можно объяснить высвобождением наземного и подземного пространства в результате снижения под кроной Angelica sylvestris покрытия видов из 1 группы – эдификаторов (Filipendula ulmiа, Alopecurus pratensis, Deschampsia caespitosa), которые формируя мощную дернину, подавляют развитие Poa nemoralis, Veronica chamaedrys, Carex nigra, Geum rivale. Сам вид Angelica sylvestris имеет стержнекорневую систему, не образует плотной дернины, и тем самым не препятствует росту патиентных видов, которые не проявляют негативной реакции на терпеноиды дудника. Кроме того, дудник, в отличии от других видов лугового травостоя, пик развития которых приходится на июнь – начало июля, наращивает основную биомассу и переходит к фазе цветения во второй половине лета (конец июля - август) и не препятствует их развитию. Можно предположить, что дудник, подавляя развитие видов-эдификаторов, благоприятствует некоторым видам с патиентным типом стратегии.
Рисунок 3. Изменение проективного покрытия Veronica chamaedrys лугового вида при увеличении покрытия Angelica sylvestris
- 3 группа представлена таким лесным видом как Melampyrum pratense.
Лесной вид Melampyrum pratense чувствителен только к высоким значениям проективного покрытия Angelica sylvestris, проективное покрытие от 3 до 55% дудника не оказывает на Melampyrum pratense видимого отрицательного влияния (рис. 4).
Этот вид является лесным и в соответствии с экологической шкалой Цыганова является семигелиофитом, т. е. способен выдерживать слабое затенение, которое формируется при средних показателях проективного покрытия Angelica sylvestris.
В ходе исследований нами была поставлена задача сравнить эдификаторную роль синузий Angelica sylvestris и эдификаторную роль отдельно растущих особей Angelica sylvestris. Всего для синузий нами было выделено 80 пробных площадок. В ходе описания морфологического строения дудника выяснилось, что растения, растущие внутри синузии, по некоторым количественным показателям достоверно отличаются от отдельно растущих особей (табл.2).
Рисунок 4. Изменение проективного покрытия вида лесной группы при увеличении проективного покрытия Angelica sylvestris
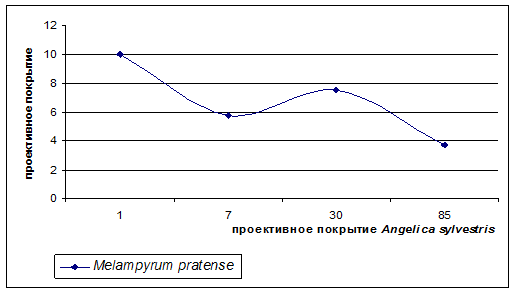
Таблица 2. Морфометрические показатели особей Angelica sylvestris, растущих отдельно и в синузии
Пло щадь, м. кв. | число особей | уровень нижнего листа, см | число листьев | длина листовой пластинки среднего нижнего листа, см | высота соцветия, см | высота расте- ния, см | |
синузия | 3,60± 0,71 | 7,30 ± 1,02 | 30,20 ± 1,31 | 3,50 ± 0,45 | 37,80 ± 1,46 | 22,20 ± 4,17 | 81,00 ± 9,87 |
отдельное растение | - | - | 20,40± 2,18 | 2,80± 0,17 | 28,40±2,05 | 34,40± 2,05 | 87,0± 5,49 |
Примечание: жирным шрифтом выделены достоверно различающиеся средние.
Достоверные отличия были получены по следующим параметрам: уровень крепления нижнего листа, длина листовой пластинки среднего и нижнего листа и высота соцветия. Полученные результаты объясняются самозатенением особей внутри синузии, которое проявляется в этилизации и удлинении вегетативных органов, при этом структурные показатели, такие как количество листьев, достоверно не изменяется. Мы предположили, что влияние синузии, на напочвенный покров отличается от влияния отдельно растущих особей Angelica sylvestris.
Анализ ценотического воздействия отдельно растущих особей показал, что Angelica sylvestris положительно взаимодействуют, увеличивая проективное покрытие, с такими видами как Geranium palustre, Carex nigra, (табл.3). Синузии Angelica sylvestris также дают положительные коэффициенты корреляции с этими видами, а так же с Galium palustre и G. uliginosum. Кроме этого было зафиксировано, что синузии Angelica sylvestris проявляют некоторое отличие от отдельно растущих особей - они демонстрируют положительную корреляцию с проростками древесных растений: березы (Betula pendula), сосны и дуба. По-видимому, синузии дудника создают несколько более благоприятные условия для проростков, затеняя и, вероятно, создавая более увлажненный микроклимат, чем крона отдельных особей.
В целом значительных различий при оценке ценотической роли отдельных растений и синузий дудника выявить не удалось.
По предварительным результатам исследований можно сделать следующие выводы:
1. В изученных сообществах Angelica sylvestris проявляет эдификаторные свойства, т. к. в зоне его воздействия снижается освещенность и изменяются проективные покрытия ряда видов.
2. Виды, имеющие разные типы жизненных стратегий, отличаются реакцией на увеличение обилия Angelica sylvestris
- виолентные виды, такие как, Filipendula ulmaria, Deschampsia caespitosa, Alopecurus pratensis, Anthriscus sylvestris, снижают свое проективное покрытие при увеличении обилия Angelica sylvestris;
- виды, чаще проявляющие патиентность (Ranunculus repens, Galium uliginosum, Geranium palustre, Molinia coerulea, Lysimachia vulgaris), не изменяют проективного покрытия с ростом обилия Angelica sylvestris.
Особи Angelica sylvestris, растущие отдельно и образующие синузию, сходны по ценотической роли в сообществе. Показано благоприятное воздействие краевых зон синузий Angelica sylvestris на всходы древесных пород.
2. Изучение роли ольхи серой в зарастании лугов
Помимо изучения эдификаторной роли некоторых видов луговых фитоценозов, провоцирующих формирование мозаичной структуры суходольных лугов, большой внимание было уделено изучению роли мелколиственных древесных пород (осины и ольхи серой) в зарастании лугов.
Ольха серая (Alnus incana) – крупный кустарник или небольшое дерево, редко дорастающее до высоты 20м. Это пионерная порода, умеренно требовательная к влаге и почве. Сероольховые леса формируются на месте заброшенных пашен, пастбищ и сенокосов, а также на вырубках, периодически затопляемых участках пойм и по окрайкам болот. Чаще всего ольха серая участвует в формировании древостоев на переувлажненных участках и участках с неустойчивым водным режимом.
Цель данной работы – изучение роли ольхи в зарастании луга. Для разрешения этой цели были поставлены следующие задачи:
1. Исследовать влияние ольхи разного возраста зарастания на изменение биотопических показателей (условия освещения, влажности, температурного режима, характеристики почвы и подстилки).
2. Исследовать влияние ольхи на живой напочвенный покров через оценку видового состава и обилие травостоя.
Нами были исследованы сероольшанниковые синузии в урочище Лахта, всего 4 синузии. При этом в каждой синузии были заложены и изучены пробные площади на участках с разным возрастом деревьев: лет, 2. –1лет, 3. – 8 – 10 лет и 4. - на примыкающем лугу. На каждом участке описывался древостой: фиксировался породный состав, возраст, высота, диаметр ствола деревьев и плотность на 1 га. Так же учитывался подлесок: фиксировался видовой состав и снимались морфометрические показатели. Для характеристики живого напочвенного покрова закладывалось по 30 учетных площадок (0,1 м2) , на которых определялся видовой состав и проективное покрытие травяного и мохового ярусов. Кроме того, на каждом участке были взяты по одному модельному укосу с площади 0,1 м2 . При изучении биотопа на всех исследуемых участках были оценены следующие показатели: уровень освещенности, c помощью сквозистомера (Ипатов и др., 1971), люксметра Ю116, цифрового фотоаппарата; температура и влажность воздуха, подстилки и верхних горизонтов почвы (глубина – 7 см) с использованием термогигрометра ТКА-ПКМ; определялась микробиологическая активность почвы методом аппликации с использованием кинопленки; содержание древесных и травянистых корней в единице объема почвы; кислотность почвы с помощью рН-метра; влажность почвы; состав и обилие опада.
Результаты исследования:
Характеристика плотности древостоя
В ряду увеличения возраста зарастания плотность ольхи снижается. При этом снижается и доля участия подлеска: в тридцатипятилетнем ольшанике мы встречаем лишь несколько особей ивы козьей (Salix caprea); на участках двадцати и десятилетнего ольшаника часто встречаются молодые особи черемухи, высотой от 1,5 до 3м. В подлеске десятилетней ольхи особенно много проростков черемухи. Древостой ольхи здесь еще не сформирован. Высота большинства особей не превышает 1,5 м и их плотность достигает 7500 шт./га.
Важным показателем структуры древостоя является уровень освещенности под его пологом. Данные измерения освещенности тремя методами (при помощи цифрового фотоаппарата, люксметра и сквозистомера) привели к сходным результатам и достоверных отличий не обнаружили. В целом, уровень освещенности увеличивается при движении к участку самого молодого возраста зарастания и на лугу достигает 95%.
В ходе зарастания луга ольхой происходят изменения температуры и влажности (табл. 3). Все замеры, необходимые для изучения температурного режима и режима увлажнения производились во второй половине дня в условиях равномерной облачности в течении 40 минут. Полученные данные показали, что температура почвы на всех уровнях (над подстилкой, в подстилке) на лугу выше, чем под пологом ольхи. Температура воздуха, напротив, ниже на лугу. Возможно, это связано с тем, что воздух под пологом древостоя медленнее остывает.
В результате измерения влажности с использованием термогигрометра нами было установлено, что подстилка и почва влажнее под пологом ольхи, в то время как на лугу влажность воздуха и на подстилке выше, чем в ольшанике. Это, вероятно, связано с обильным живым напочвенным покровом луга. Здесь его среднее проективное покрытие составляет 85%, тогда как в тридцатипятилетнем ольшанике – всего 10%.
Для исследования почвы на участке каждого возраста зарастания ольхи и на лугу были сделаны почвенные разрезы и прикопки. На их основе мы характеризовали данный тип почв как дерново-подзолистые, по гранулометрическому составу – средний суглинок. В разрезах всех участков с глубины 15 см и ниже обнаруживаются рыжие включения железа – свидетельство режима переменного увлажнения. В большом количестве встречаются ходы дождевых червей, муравейники. По мере увеличения возраста зарастания уменьшается горизонт дерновины. На лугу он достигает 5 см, в десятилетней ольхе – 2-3см, а под пологом тридцатипятилетней ольхи отсутствует вовсе. Мощность гумусового горизонта в этом ряду увеличивается с 1 – 2 см на лугу до 4 см в почве под тридцатипятилетней ольхой.
Таблица 3. Характеристика температуры и влажности.
Участок | Показания термометра | Показания гигрометра | ||||||
воздух | над подстилкой | Подстилка | почва | воздух | над подст. | подстилка | почва | |
30 -35 лет | 23.1 ± 0.3 | 23.06 ± 0.17 | 21.72 ± 1.1 | 20.18 ± 0.4 | 60.8 ± 0.26 | 73.22 ± 3.18 | 87.1 ± 1.73 | 95.4 ± 3.99 |
1лет | 23.14 ± 0.65 | 23.56 ± 0.15 | 21.82 ± 1.0 | 21.62 ± 0.7 | 58.16 ± 0.85 | 71.6 ± 1.92 | 83.66 ± 4.15 | 93.52 ± 3.17 |
лет | 22.86 ± 0.69 | 24.72 ± 0.19 | 22.72 ± 0.7 | 22.42 ± 0.5 | 63.18 ± 3.87 | 77.52 ± 4.04 | 84.7 ± 1.80 | 89.34 ± 3.80 |
луг | 21.94 ± 0.9 | 25,2 ± 0.33 | 25.86 ± 0.9 | 24.16 ± 4.8 | 66.86 ± 3.29 | 80.72 ± 4.58 | 79.38 ± 1.44 | 79.74 ± 3.64 |
Рисунок 5. Состав подстилки Рисунок 6. Содержание корней в почве
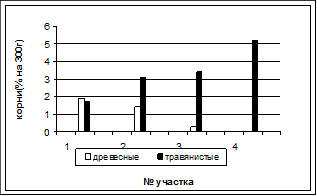
 Также по мере зарастания луга изменяется состав подстилки и содержание корней в почве (рис. 5). После разбора на фракции выявилась тенденция к снижению лиственного опада и веток в ее составе по направлению к лугу. При этом содержание остатков травянистых растений разной степени разложения увеличивается.
Также по мере зарастания луга изменяется состав подстилки и содержание корней в почве (рис. 5). После разбора на фракции выявилась тенденция к снижению лиственного опада и веток в ее составе по направлению к лугу. При этом содержание остатков травянистых растений разной степени разложения увеличивается.
Таблица 4. Характеристика кислотности почвы.
Участок | Ср. знач. рН | ошибка ср. знач | t-крит |
30-35 лет | 5,31 | 0.03 | 0.02 |
18-20 лет | 5,27 | 0.04 | 0.01 |
8-10 лет | 5,53 | 0.09 | 0.01 |
Луг | 5,84 | 0.03 | 0.01 |
При обработке данных кислотности почвы на участках разного возраста ольхи и на лугу использовался критерий Стьюдента. При взятом уровне достоверности 0,05 значимых различий не обнаружено. Почва на всех участках слабокислая (табл. 4).
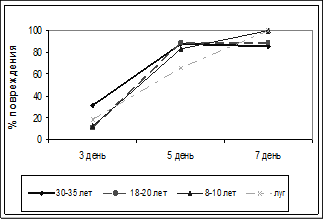 Рисунок 7. Микробиологическая активность почвы.
Рисунок 7. Микробиологическая активность почвы.
Также была проведена оценка микробиологической активности почвы методом аппликации кинопленок. Метод аппликации дает высокую степень корреляции с нитрогеназной активностью и рестриктазным методом, применяемым для оценки состава и количества микробиоты, поэтому по интенсивности повреждения эмульсионного слоя пленки мы можем косвенно судить об интенсивности деятельности почвенной микробиоты. Достоверных отличий в скорости повреждения эмульсионного слоя пленки на участках разного возраста зарастания выявлено не было (рис. 7), что нельзя сказать о развитии растительного напочвенного покрова.
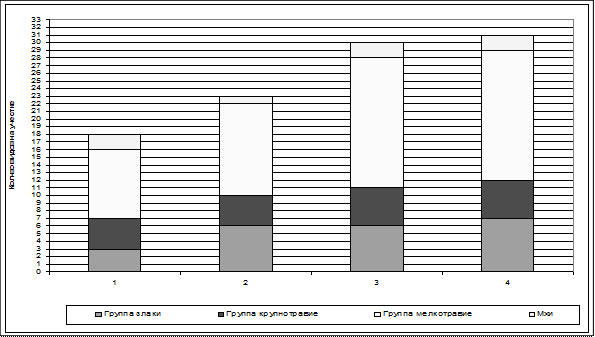
При анализе видового и ценотического состава установлена следующая тенденция: в целом, по мере зарастании луга ольхой снижается доля лугового мелкотравья: Veronica chamaedys, Galium borealis, Vicia cracca и др. Также снижается доля участия в луговых злаков, таких как Alopecurus pratensis, Phleum pratensis, Festuca rubra . Под пологом ольхи сохраняются только наиболее теневыносливые злаки (Elitrigia repens). При этом крупнотравье: дудник (Angelica sylvestris ), купырь (Anthriscus sylvestris ) и таволга (Filipendula ulmaria) обнаруживается на всех участках (рис. 8, 9).
Рисунок 8. Увеличение разнообразия видов в растительном покрове по мере снижения возраста ольхи серой.
При зарастании луга ольхой серой, как и при зарастании осиной, увеличивается сомкнутость и плотность древостоя, что сопровождается изменением биотопа, и в первую очередь проявляется в изменении условий освещенности. Изменяются показатели температуры и влажности: под пологом ольхи температура воздуха, подстилки, ее поверхности и почвы ниже, чем на лугу, а влажность воздуха, подстилки, ее поверхности и почвы под пологом, наоборот, выше, чем на лугу. Зарастание ольхой влияет на состав подстилки: возрастает доля листового опада, древесных остатков, из-за снижения общего проективного покрытия трава снижается доля ветоши и постепенно исчезает дернина, на фоне увеличения гумусового горизонта. В живом напочвенном покрове сокращается обилие и разнообразие луговых видов, но и лесные виды не проявляют еще своего активного участие. Сохраняют свою позицию только некоторые крупные многолетние травы: дудник, таволга, возрастает участие рудеральных видов, например, крапивы.
По мере зарастания луга осиной в живом напочвенном покрове возрастает роль древесных растений – увеличивается их фитомасса и видовое разнообразие. В ходе сукцессии, осины дифференцируются, снижается плотность их древостоя, но возрастает участие других пород (береза, ель). Изменяется травянистый состав живого напочвенного покрова. Под пологом осины увеличивается роль лесных видов, изменяется состав граминоидов, луговые виды сменяются на лесные виды. Так же в живом напочвенном покрове снижается доля участия лугового крупнотравья.
Это первоначальные результаты исследования, которые будут корректироваться и дополняться в процессе дальнейшей работы. Всего за время экспедиции 2009 нашей группой было сделано 14 полных геоботанических описаний и описано 620 учетных площадок, проведен пофракционный и весовой анализ 24 модельных укосов, проанализировано 4 полных почвенных разреза и 10 прикопок.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 |
