

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
РАЗРАБОТКА ИММУНОФЕРМЕНТНОЙ ТЕСТ-СИСТЕМЫ ДЛЯ ВЫЯВЛЕНИЯ СТАФИЛОКОККОВОГО СЕПСИСА
1, 1, 2, 3, 2, 1
1Институт молекулярной биологии и биохимии им. М.А. Айтхожина, Алма-Ата, Казахстан; 2ГУ НИИ биохимии СО РАМН, Новосибирск, Россия; 3Институт химической биологии и фундаментальной медицины СО РАН, Новосибирск.
Контантный электронный адрес: *****@***com
Золотистый стафилококк вызывает различные инфекции у человека, включая эндокардиты, остеомиелиты, артриты, раневой сепсис, кожные абсцессы, маститы, кератиты и является причиной более 40% случаев госпитальных инфекций. Одним из основных факторов патогенности золотистого стафилококка является альфа-токсин (САТ), обладающий рядом активностей: цитолитической, иммуномодулирующей, дермонекротической и летальной [1]. Септические состояния стафилококкового происхождения сопровождаются появлением в крови САТ, который используют в качестве этиологического маркера сепсиса. Между тем, в настоящее время в странах как СНГ, так и дальнего зарубежья отсутствуют коммерческие тест-системы, позволяющие надежно и быстро определить этиологическую природу сепсиса. Биотехнологический подход к решению проблемы диагностики стафилококкового сепсиса заключается в получении рекомбинантного САТ и моноклональных антител, полученных к различным его эпитопам, и конструировании иммуноферментных тест-систем на их основе.
Целью работы явилось получение моноклональных антител к нативной молекуле САТ, рекомбинантной формы САТ и разработка ингибиторной иммуноферментной тест-системы на их основе.
Методы исследования
Получение рекомбинантного САТ
В работе использовали следующие бактериальные штаммы и векторные ДНК. RecA(+) штаммы Escherichia coli: C600 и BL21(DE3), а также recA(-) штаммы – DH-5α, XL-Blue, XL-blue2, JM 109. Штаммы Staphylococcus aureus: Wood 46, O15. Векторные ДНК: pBluescript II SK (+), pBR322 («Stratagene», США) и сконструированный в лаборатории вектор pREB-6His для прямой экспрессии генов в клетках E. coli на основе pBR322, содержащей регуляторную область гена recA Proteus mirabilis.
Выбор олигонуклеотидов-праймеров проводили с помощью программы OLIGO (версия 3.3). Использовали следующие нуклеотидные последовательности праймеров для амплификации структурной области гена hla S.aureus
PR22F: 5'- GGAATTCATGAAAACACGTATAGTC-3'
PR23R: 5' AGGATCCTTAATTTGTCATTTCTTCTT 3'
PR31R: 5' AGGTCACCATTTGTCATTTCTTCTTTTTC 3'
Праймер PR22F (прямой) и обратные праймеры PR23R и PR31R предназначены для амплификации и последующей встройки структурной области гена hla S.aureus в экспрессирующий вектор pREB-6His по сайтам EcoRI (PR22F) и BamHI (PR23R) или BstEII (PR31R). В случае применения праймера PR31R обеспечивалась состыковка гена с нуклеотидной последовательностью вектора, кодирующей 6-гистидиновую АК-последовательность в С-концевой области экспрессируемого белка. Продукты амплификации анализировали с помощью электрофореза в 5-12% ПААГ или в 2% агарозном геле.
Процедуру лигирования фрагмента ДНК с плазмидой-вектором по липким концам проводили согласно инструкции фирмы-изготовителя ДНК-лигазы фага Т4. После завершения процесса лигирования аликвоты реакционной смеси после предварительного диализа использовали для трансформации компетентных клеток E. coli методом электропорации. Уровень трансформации в среднем составлял 109 колоний на 1 мкг суперскрученной ДНК плазмиды pBR322.
Колонии E. coli соскребали, ресуспедировали в ТЕ. Микробные клетки лизировали 0,2 М NaOH в присутствии 2% SDS. К лизату добавляли KCl, сахарозу и бромфеноловый синий, перемешивали и осаждали хромосомную ДНК центрифугированием при 10000 g в течение 20 мин. Супернатант, содержащий плазмидную ДНК, анализировали электрофорезом в агарозном геле.
Колонии E.coli, содержащие рекомбинантные плазмиды, пересевали на LB-среду. В культуру вносили налидиксовую кислоту до концентрации 50 мкг/мл в качестве индуктора, активирующего ген recA, и продолжали выращивание клеток еще 3-5 ч. Затем клетки осаждали при 5000 g в течение 10 мин. Осадок клеток лизировали кипячением в трис-буфере в присутствии 100 мМ DTT, 2% SDS, 0,1% BPB и 10% глицерина. Лизат анализировали электрофорезом в ПААГ в денатурирующих условиях по методу Лэммли.
Препаративную наработку рекомбинантного САТ проводили по аналогичной схеме в 100–500 мл LB-среды. Разрушение бактериальных клеток проводили либо путем многократной заморозки-разморозки, либо при помощи ультразвука. В первом варианте собранные центрифугированием клетки E.coli ресуспендировали в 0,1 М Трис-HCl буфере при pH 6,8, содержащем 0,01% Тритон Х-100 и лизоцим (5 мг/мл). Суспензию подвергали многократному (8-10 раз) замораживанию-оттаиванию. ДНК разрушали ДНК-азой I. Лизат центрифугировали при 20000 g в течение 30 мин, и рекомбинантную форму САТ, содержащего в С-концевой области 6-гистидиновую аминокислотную последовательность выделяли методом аффинной хроматографии на колонке с Ni2+-NTA-сефарозой-6В согласно инструкции фирмы-изготовителя аффинного сорбента. Определение гемолитической активности САТ проводили по методу [2].
Получение моноклональных антител (МКА) к нативной форме САТ.
Для иммунизации мышей линии BALB/c, а также для тестирования гибридом использовали 70% чистоты препарат САТ, любезно предоставленный профессором (Институт физиологии, Ташкент). По данным SDS-электрофореза САТ был гомогенен и имел кажущуюся ММ около 34 кДа. Для получения высокоактивного иммуногена и снижения токсичности САТ конъюгировали с активированной целлюлозой по методу [3]. Шесть мышей линии BALB/c иммунизировали по следующей схеме. 20 мкг САТ на целлюлозе вводили внутрибрюшинно, трехкратно с интервалом 2 недели. Бустерную инъекцию проводили через 2 недели по 7 мкг чистого САТ в физиологическом растворе в хвостовую вену. Слияние клеток проводили на 3-й день по стандартному методу [4] с небольшими модификациями. Для культивирования клеток использовали среду RPMI-1640, содержащую 20% фетальной телячьей сыворотки и антибиотики. Клонирование гибридом проводили трижды методом лимитирующих разведений. Наработку антител стабильно продуцирующим моноклоном производили in vitro. Антитела осаждали из культуральной среды насыщенным раствором сульфата аммония с последующим диализом против фосфатного буферного раствора. Для скрининга гибридом, анализа и разработки тест-системы были использованы различные варианты иммуноферментного анализа [10]. Мышиные иммуноглобулины выявляли пероксидазным конъюгатом "кролик-анти-мышь" (БИОС, Новосибирск) или "козел-анти-мышь" (BioRad, США). Для определения субкласса полученных МКА использовали антиизотипические сыворотки ( Sigma, США).
Результаты и обсуждение
Рекомбинантный стафилококковый альфа-токсин. Для получения рСАТ структурную область гена САТ S. aureus шт. Wood 46 клонировали в клетках E. coli в составе сконструированного нами экспрессирующего вектора pREB (рис.1), предназначенного для прямой экспрессии генов прокариот под контролем регуляторной области гена recA Proteus mirabilis. Было проведено два варианта встройки нуклеотидной последовательности, кодирующей полипептид-предшественник зрелого САТ. Первый вариант предусматривал встройку гена по сайтам EcoRI и BamHI, в результате которой должен был синтезироваться предшественник САТ без дополнительных аминокислотных остатков как с N-, так и с С-концов молекулы. По второму варианту встройки, осуществляемому по сайтам EcoRI и BstEII, предшественник САТ должен был содержать в С-концевой области 6-гистидиновую аминокислотную последовательность, предназначенную для аффинной очистки рекомбинантного САТ. При индукции трансформированных клеток E. coli налидиксовой кислотой или митомицином С наблюдался высокий уровень синтеза двух рекомбинантных форм САТ (~30% от суммарного клеточного белка), причем токсин эффективно секретировался в периплазму и, что следует особо отметить, в культуральную среду. Синтезированный рСАТ обладал высокой гемолитической активностью, сравнимой с таковой у нативного САТ из S. aureus. Этот факт свидетельствует о корректном процессинге чужеродного для E. coli САТ, являющегося типичным секреторным белком грамм-положительных бактерий. Эффективная экспрессия гена САТ в клетках E. coli, сопровождаемая секрецией токсина в периплазму и культуральную среду, обусловлена, по-видимому, использованием в векторе регуляторной области гена RecA, который, как известно, играет основную роль в активации SOS-ответа, включение которого сопровождается активацией синтеза ряда секреторных белков.
Была исследована также гемолитическая активность и локализация в клетке рекомбинантной формы САТ, содержащей дополнительную 6-гистидиновую аминокислотную последовательность на С-конце молекулы. Как и в первом случае, рекомбинантный белок обладал гемолитической активностью и секретировался в периплазму и культуральную жидкость рСАТ, содержащий дополнительную С-концевую 6-гистидиновую последовательность, очищали аффинной хроматографией на Ni2+-NTA-сефарозе-6В и использовали для конструирования диагностической тест-системы на основе ингибиторного ИФА.
Моноклональные антитела к САТ. Мышиные МКА получали с помощью стандартной гибридомной технологии. В результате слияния спленоцитов и клеток из региональных лимфоузлов с миеломной клеточной линией Х63.Ag8.653 получили высокий выход гибридом. Эффективность гибридизации (процент ячеек с гибридомами) составила свыше 90%, а специфическая эффективность - 6% (процент антителопродуцирующих гибридом от общего их количества). После первого скрининга 11 гибридом подвергли криоконсервации. Однако после разморозки и последующего культивирования у гибридом наблюдалось уменьшение продукции антител вплоть до их полного исчезновения. Только одна гибридома D3 показала при длительном культивирование стабильную антителопродукцию и хорошие ростовые свойства. Данную гибридому клонировали 5 раз, затем подвергли препаративному накоплению в культуральной среде. При этом продукция моноклональных антител гибридомой D3 при длительном культивировании в культуральной среде, определяемая в непрямом ИФА, достигала титра 1:78000.
Ранее мы получали МКА к САТ при использовании метода иммунизации in vitro [6]. Однако эти антитела имели IgM-изотип и показывали высокую полиспецифичность. Кроме того, полученные IgM-продуцирующие гибридомы оказались не стабильными и через несколько пассажей потеряли способность к антителопродукции.
Изотипический анализ показал, что секретируемые МКА D3 относятся к IgG1- подклассу (рис. 2).
При оценке специфичности МКА D3 в непрямом ИФА в качестве гетерологичного антигена использовали стафилококковый энтеротоксин А (СЭА, производства НИВС, УФА). Как видно из рис. 3, МКА D3 показывали высокую специфичность к САТ, что также было подтверждено в иммуноблоттинге (данные не показаны).
Связывание МКА D3 с рекомбинантным САТ мы определяли в непрямом и ингибиторном варианте ИФА. Непрямой ИФА мы провели в различных условиях сорбции рСАТ (в нейтральном и щелочном буферном растворе). Как видно из рис. 4, МКА D3 показывали связывание с рСАТ, иммобилизованным на планшете как при нейтральном, так и при щелочном рН, однако щелочной рН предпочтительнее (карбонат-бикарбонатный буферный раствор, рН 9,1).
Сущность ингибиторного варианта ИФА заключается в способности свободного рСАТ подавлять связывание МКА с иммобилизованным рСАТ. В предварительных экспериментах была определена оптимальная концентрация рСАТ для иммобилизации (2-4 мкг/мл) и рабочее разведение МКА D3, соответствующее 0,35 мкг/мл. При проведении ингибиторного ИФА свободный рСАТ в различных концентрациях инкубировали совместно с МКА D3 в ячейках с иммобилизованным рСАТ. После двухчасовой инкубации связавшиеся антитела выявляли антивидовым конъюгатом. По степени торможения реакции связывания судили о наличии рСАТ в растворе. Как видно из рис. 5, чувствительность данной системы составила 0,15 - 0,30 мкг/мл.
Таким образом, полученный рекомбинантный САТ по функциональным и иммунохимическим свойствам не отличался от нативного аналога. Моноклональные антитела D3, специфичные к нативному САТ, оказались способными выявлять рекомбинантный САТ как фиксированный на полистирольных планшетах, так и в жидкой фазе в ингибиторном ИФА. Ингибиторный ИФА на основе моноклональных антител D3 к САТ и рекомбинантного САТ позволяет выявлять САТ в жидких средах в концентрациях до 0,30 мкг/мл. Предполагается использование разработанной тест-системы для оценки продукции САТ токсигенными штаммами стафилококка в микробиологических культурах, а также определения содержания САТ в крови у септических больных в диагностических целях.
Литература
1. Bhakdi S., Tranum-Jensen J. Alpha-toxin of Staphylococcus aureus//Microbiol. Rev.-1991.-V.55.-N4.-P.733-751
2. Bernheimer A. W. Assay of Hemolytic toxins//Methods in Enzymology.–1988.-V.165.–P.213-217
3. , Гурвич высокоемкого иммуносорбента на основе суспензии целлюлозы//Бюлл. экспер. биол.-1981.-№7.-с.68-70.
4. Campbel A. Monoclonal antibody technology. Amsterdam, Elsevier,1984,.265 p.
5. Иммуноферментный анализ. Антитела. Методы. Кн. 2, М, 1991.- с.152-238.
6. , К, , Навасардянц моноклональных антител к стафилококковому a-токсину при использовании техники иммунизации in vitro//Бюлл. экспер. биол.-1991.-№7.-с.73-75.
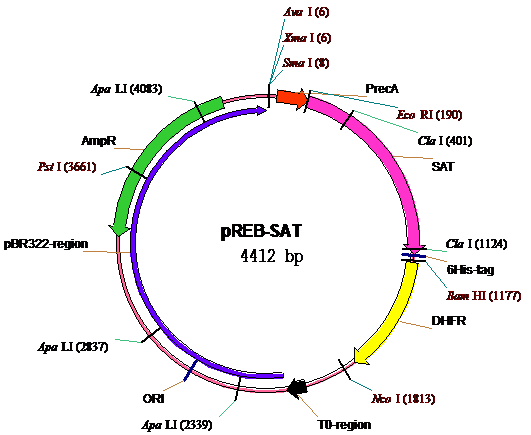 |
Рис. 1. Схема рекомбинантной плазмиды pREB-SAT, обеспечивающей индуцируемую прямую экспрессию структурной области гена стафилококкового альфа-токсина S.aureus шт. Wood 46 под контролем регуляторной области гена recA Proteus mirabilis
Примечание:
- PrecA – регуляторная область гена recA Proteus mirabilis, включающая recA-промотор, SOS-box и участок SD;
- SAT – структурная область гена hla S.aureus, кодирующая полипептид-предшественник альфа-токсина;
- DHFR - структурная область гена дигидрофолатредуктазы;
- T0-region - участок плазмиды, содержащий терминатор транскрипции t0 бактериофага лямбда;
- AmpR - структурная область гена бета-лактамазы
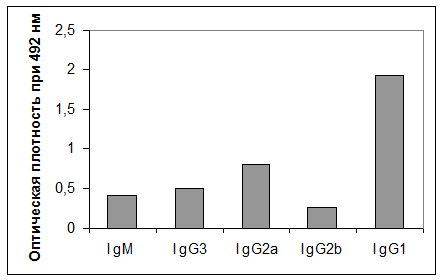 |
Рис. 2. Изотипический анализ МКА D3
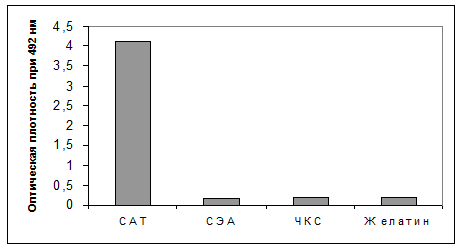 |
Рис. 3. Специфичность связывания МКА D3 в непрямом ИФА
Обозначения: СЭА - стафилококковый энтеротоксин А, ЧКС - чистая культуральная среда
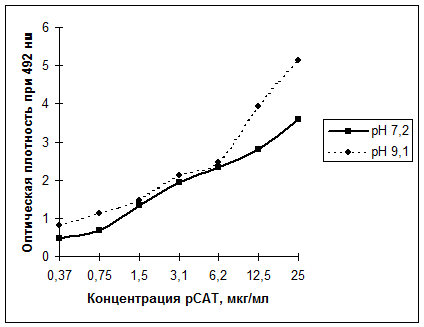 |
Рис. 4. Связывание МКА D3 с рекомбинантным САТ, иммобилизованным на полистироловом планшете при различном рН буферного раствора.
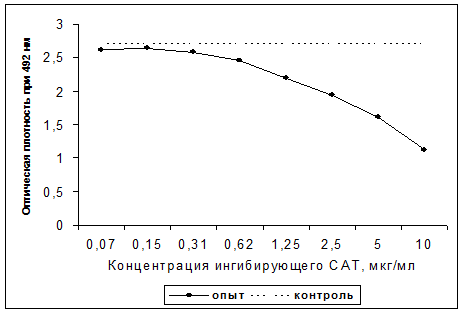 |
Рис. 5. Титрование рСАТ в ингибиторном ИФА
Примечание: контроль - прямое связывание МКА с фиксированным на планшете рСАТ, опыт - ингибирование связывания МКА с фиксированным на планшете рСАТ растворимым в в жидкой фазе нативным САТ.
