
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Задача 1.21. Исследовали кинетику окисления L-лактата лактатдегидрогеназой в водно-солевых экстрактах двух органов (печени и сердца), взятых у одного животного. Перед опытами тканевые экстракты диализовали против воды.
В 1см кювету спектрофотометра добавляли 0,5 мл экстракта, 0,3 мл 10 мМ раствора NAD+, 0,2 мл 30 мМ раствора семикарбазида и 1,0 мл натрий-фосфатного буфера, рН 7,6. Затем в кювету добавляли 100 мкМ раствор L-лактат натрия в количествах, указанных в таб. 21, и водой доводили объем пробы в каждой кювете до 3 мл. После добавления лактата содержимое кюветы тщательно перемешивали и измеряли через каждые 30 с оптическую плотность пробы на спектрофотометре при длине волны 340 нм. Полученные данные приведены в табл. 1.21.
Таблица 1.21
Время. мин | Оптическая плотность при 340 нм | |||||||
Объем добавленного L-лактата, мл 100 мМ раствора | ||||||||
0,2 | 0,25 | 0,5 | 1,0 | 0,2 | 0,25 | 0,5 | 1,0 | |
Экстракт печени | Экстракт сердца | |||||||
0 | 0,016 | 0,012 | 0,023 | 0,031 | 0,013 | 0,030 | 0,023 | 0,015 |
0,5 | 0,050 | 0,052 | 0,080 | 0,105 | 0,052 | 0,074 | 0,082 | 0,084 |
1,0 | 0,084 | 0,092 | 0,137 | 0,174 | 0,090 | 0,119 | 0,137 | 0,155 |
1,5 | 0,117 | 0,131 | 0,208 | 0,252 | 0,127 | 0,162 | 0,196 | 0,222 |
2,0 | 0,149 | 0,169 | 0,249 | 0,328 | 0,163 | 0,204 | 0,258 | 0,290 |
2,5 | 0,180 | 0,207 | 0,305 | 0,397 | 0,199 | 0,245 | 0,315 | 0,350 |
Необходимо: 1) построить кинетические кривые реакции (графики зависимости оптической плотности от времени), провести касательные к начальным участкам кривых и определить начальную скорость реакции, катализируемой ЛДГ; 2) используя полученные значения начальных скоростей, построить графики зависимости v0 от [S]; 3) линеаризовать графики v0 от [S] для определения величин Км и Vmax; сравнить кинетические характеристики ЛДГ в экстрактах печени и сердца, дать обоснование полученным данным.
Задача 1.22. Гомогенат сердца кролика, содержащий малатдегидрогеназу, способен окислять в присутствии NAD+ cледующие субстраты: L-малат, мезотартрат и DL-оксибутират.
А. Окисление малата. В 4-см спектрофотометрическую кювету добавляли 4 мл 0,3 М глицинового буфера, рН 10, и 0, 4 мл 50 мМ раствора NAD+. Затем в кювету добавляли различные объемы 5 мМ раствора L-малата натрия и общий объем пробы доводили водой до 11,9 мл. Реакция начинали добавлением 0,1 мл разбавленного в 10 раз экстракта сердечной мышцы и измеряли оптическую плотность при 340 нм через каждые 30 с в течение 3 мин. Полученные результаты приведены в табл. 1.22А.
Таблица 1.22А
Время, мин | Оптическая плотность при 340 нм | |||||
Объем добавленного 5 мМ раствора L-ьалата, мл | ||||||
2,4 | 1,2 | 0,72 | 0,48 | 0,24 | 0,12 | |
0,5 1,0 1,5 2,0 2,5 3,0 | 0,085 0,162 0,236 0,310 0,385 0,458 | 0,060 0,123 0,184 0,245 0,395 9,367 | 0,070 0,120 9,169 0,220 0,269 0,319 | 0,048 0,086 0,123 0,160 0,199 0,236 | 0,027 0,051 0,074 0,099 0,122 0,145 | 0,019 0,033 0,046 0,061 0,074 0,087 |
Б. Окисление мезотартрата. Условия опыта для изучения окисления трантрата были такими же, как и в случае малата, но вследствие того, что скорость реакции в этом случае была значительно меньше, использовали большие количества экстракта. Все добавки были такими же, как и при изучении окисления малата, за исключениями: 1) были изменены концентрации субстрата (указаны в табл. 1.22Б) и 2) объем пробы доводили водой до 11,5 мл и реакцию начинали добавлением 0,5 мл неразбавленного экстракта. Полученные данные приведены в табл.22.2
Таблица 1.22Б
Время, мин | Оптическая плотность при 340 нм | |||||
Объем добавленного 10 мМ раствора мезотартрата, мл | ||||||
2,4 | 1,2 | 0,6 | 0,36 | 0,28 | 0,18 | |
0,5 1,0 1,5 2,0 2,5 3,0 | 0,034 0,068 0,100 0,133 0,166 0,198 | 0,029 0,054 0,078 0,102 0,127 9,152 | 0,024 0,040 9,057 0,073 0,090 0,106 | 0,012 0,023 0,035 0,046 0,056 0,067 | 0,008 0,018 0,027 0,036 0,045 0,055 | 0,008 0,015 0,022 0,028 0,035 0,042 |
В. Окисление DL-оксибутирата. Этот субстрат окисляется со скоростью, промежуточной между скоростями окисления малата и мезотартрата. Поэтому при измерении скорости реакции брали разные объемы субстрата (указаны в таблтице), конечный объем пробы доводили водой до 11, 9 мл и реакцию начинали добавлением 0,1 мл неразбавленного экстракта. Полученные данные приведены в табл. 1.22В
Таблица 1.22В
Время, мин | Оптическая плотность при 340 нм | |||||
Объем добавленного 50 мМ раствора оксибутирата, мл | ||||||
2,4 | 1,2 | 0,72 | 0,48 | 0,36 | 0,24 | |
0,5 1,0 1,5 2,0 2,5 3,0 | 0,068 0,117 0,164 0,213 0,259 0,307 | 0,055 0,094 0,130 0,170 0,208 9,244 | 0,040 0,069 9,098 0,127 0,156 0,184 | 0,030 0,054 0,077 0,101 0,125 0,148 | 0,021 0,040 0,060 0,079 0,099 0,119 | 0,019 0,034 0,048 0,063 0,078 0,093 |
Задача 1.23. При действии фосфодиэстеразы на ее субстрат были получены данные, приведенные в табл. 1.23.
Таблица 1.23
[S], 10-3 М | Начальная скорость, мкмоль/мин | [S], 10-3 М | Начальная скорость, мкмоль/мин |
2,50 | 0,0212 | 0,66 | 0,0148 |
1,66 | 0,0198 | 0,50 | 0,0137 |
1,00 | 0,0176 | 0,40 | 0,0117 |
Определите величины Кm и Vmax.
Раздел 2. Ингибиторы ферментов
На активность ферментов оказывают влияние множество факторов, среди которых важную роль играют ингибиторы, воздействие которых снижает активность ферментов.
Различают обратимое и необратимое ингибирование. Необратимый ингибитор или каталитический яд, взаимодействуя с ферментом, снижает его активность до нуля.
Яркий пример необратимого ингибирования – ингибирование ацетилхолинэстеразы фторорганическими соединениями (рис. 2.1.). Ацетилхолинэстераза катализирует гидролиз ацетилхолина, избыток которого может полностью блокировать передачу нервного импульса через синапс. Поэтому фторорганические соединения являются сильными ядами.
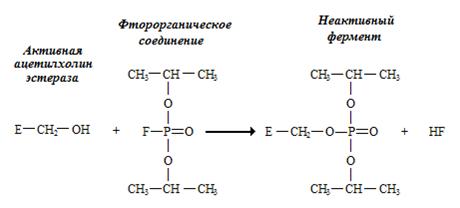
Рис. 2.1. Необратимое ингибирование ацетилхолинэстеразы
Другой пример – действие аспирина на циклооксигеназу. Ацетилсалициловая кислота ацилирует фермент в активном центре по остатку серина (рис. 2.2.).
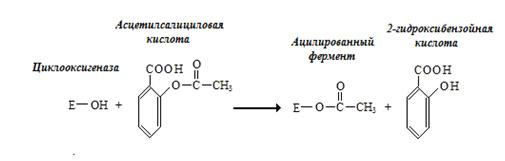
Рис. 2.2. Необратимое ингибирование циклооксигеназы
Необратимым ингибитором для ферментов, содержащих SH-группу цистеина в активном центре, является йодуксусная кислота и йодацетамид (рис. 2.3.).
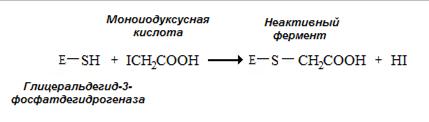
Рис. 2.3. Необратимое ингибирование глицеральдегид-3-фосфатдегидрогеназы
Обратимые ингибиторы, в отличие от необратимых образуют с ферментом динамический комплекс, отличающийся по своим кинетическим свойствам от свободного фермента.
Характерная черта обратимого ингибирования – наличие равновесия между ферментом и ингибитором. При этом константа равновесия или константа ингибирования (Кi) служит мерой сродства фермента и ингибитора и выражает эффективность действия ингибитора.
Можно выделить четыре типа обратимого ингибирования: конкурентное, неконкурентное, бесконкурентное и ингибирование смешанного типа.
Общая схема ингибирования:
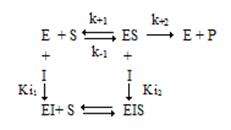
2.1. Конкурентное ингибирование
Конкурентный ингибитор – это соединение, обладающее структурным сходством с субстратом. Поэтому такой ингибитор способен взаимодействовать с активным центром фермента, конкурируя с истинным субстратом.
Схема конкурентного ингибирования
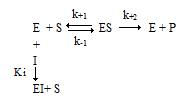
График ингибирования в координатах Лайнуивера-Берка выглядит следующим образом (рис. 2.4).
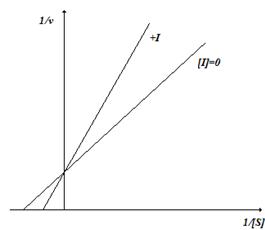
Рис. 2.4. Конкурентное ингибирование
При этом типе ингибирования увеличивается Km. Скорость реакции снижается, так как комплекс фермент-ингибитор является неактивным (не распадается с образованием продуктов). Однако максимальная скорость реакции (Vmax) не изменяется.
Уравнение Михаэлиса-Ментен в данном случае имеет вид:
![]() (1)
(1)
Где Km(каж) равна:
![]() (2)
(2)
Степень ингибирования зависит от концентраций субстрата и ингибитора, и при достаточно большой концентрации субстрата ингибирование может быть подавлено.
Пример конкурентного ингибирования – ингибирование сукцинатдегдрогеназы малоновой кислотой (рис 2.5).
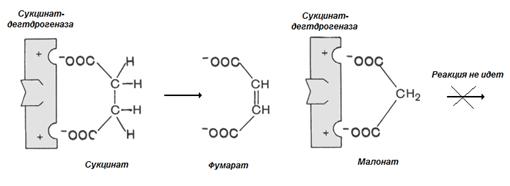
Рис. 2.5. Малонат – конкурентный ингибитор сукцинатдегидрогеназы
2.2. Неконкурентное ингибирование
В случае неконкурентного ингибирования ингибитор не оказывает влияния на взаимодействие фермента и субстрата (рис. 2.6), уменьшая скорость реакции (Vmax(каж)<Vmax).
Схема неконкурентного ингибирования
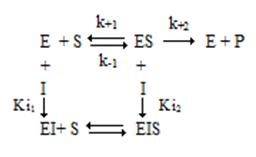
При этом Ki1=Ki2=Ki. Уравнение Михаэлиса-Ментен выглядит следующим образом:
![]() (3)
(3)
Где Vmax(каж) имеет значение:
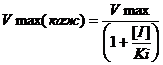 (4)
(4)
Линейный график в координатах двойных обратных величин имеет следующий вид (рис. 2.6.). Степень ингибирования зависит от концентрации ингибитора и не зависит от концентрации субстрата.
Примером неконкурентного ингибирования является ингибирование α-амилазы мальтозой (продуктом реакции). В промышленности применяется противогрибковый препарат этоний, который является неконкурентным ингибитором сахаразы грибов. В медицине его используют как антисептик.
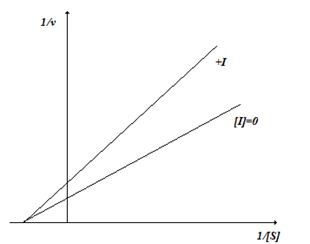
Рис. 2.6. Неконкурентное ингибирование
2.3. Бесконкурентное ингибирование
Этот тип ингибирования наблюдается в случае, когда ингибитор способен связываться исключительно с фермент-субстратным комплексом.
Схема бесконкурентного ингибирования:

При данном типе ингибирования в равной степени изменяется как константа Михаэлиса, так и максимальная скорость реакции:
![]() . (5)
. (5)
Где , а ![]() . (6)
. (6)
Так как и константа Михаэлиса и максимальная скорость изменяются в равной степени, то в координатах Лайнуивера-Берка графики имеют вид параллельных прямых (рис.2.7).
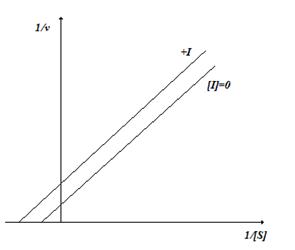
Рис. 2.7. Бесконкурентное ингибирование
Бесконкурентный тип ингибирования часто встречается в сложных полисубстратных реакциях.
2.4. Смешанный тип ингибирования
Типы ингибирования, рассмотренные выше, являются предельными случаями в широком спектре возможных эффектов. В случае двухсубстратных реакций при определенной концентрации ингибитора часто можно наблюдать ингибирование смешанного типа.
Общая схема ингибирования смешанного типа.
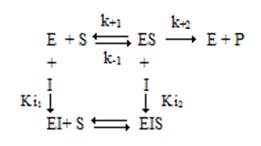
При этом Ki1 ≠ Ki2. Изменяются как константа Михаэлиса, так и максимальная скорость, но не в одинаковой степени:
![]() (7)
(7)

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 |
