
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Смешанное конкурентное-неконкурентное ингибирование. В этом варианте ингибирования Ki2˃ Ki1. Сродство фермента к субстрату в присутствии ингибитора данного типа увеличивается, а максимальная скорость ферментативной реакции снижается (рис. 2.8).
 ,
,  (8)
(8)

Рис. 2.8. Смешанное конкурентное-неконкурентное
Смешанное неконкурентное-бесконкурентное ингибирование. В этом случае Ki1 ˃ Ki2. Этот ингибитор снижает и константу Михаэлиса и максимальную скорость реакции (рис. 2.9).
 ,
,  (9)
(9)

Рис. 2.9. Смешанное неконкурентное-бесконкуоентное
Примером смешанного ингибирования является воздействие ртутьорганического соединения мертиолата на сахаразу грибов. Это вещество широко используется в промышленности для подавления роста микромицетов.
2.5. Методы определения константы ингибирования
Величины Кi можно измерить различными методами, как экспериментально, так и при помощи расчетов.
Если известна только одна концентрация ингибитора, то константу ингибирования можно определить, используя графики в координатах Лайнуивера-Берка. Найдя кажущиеся и истинные значения константы Михаэлиса и максимальной скорости для двух графиков (в отсутствии и в присутствии ингибитора) и подставив эти значения в формулы (2), (4), (7), (9), (10), рассчитывают ингибиторную константу.
Наиболее удобен для определения константы ингибирования метод Диксона. Этот простой графический метод позволяет определять Кi непосредственно. Если определять скорость реакции в условиях постоянной концентрации субстрата и различных концентраций ингибитора, то график зависимости 1/v от [I] будет представлять собой прямую (рис. 2.10.). В этом случае для определения ингибиторной константы достаточно определить скорость реакции всего для двух концентраций субстрата.

Рис. 2.10. Графическое определение ингибиторных констант (метод Диксона). Во всех случаях S1˃S2
2.6. Субстратное ингибирование
Для многих ферментативных реакций при увеличении концентрации субстрата начальная скорость ферментативной реакции проходит через максимум, а затем уменьшается. Подобного рода зависимость можно описать, исходя из предположения об образовании в процессе реакции непродуктивного тройного комплекса ЕS2. В этом случае кинетическая схема ферментативной реакции выглядит следующим образом:

Зависимость скорости реакции от концентрации субстрата после математической обработки этой схемы имеет вид:
![]() (11)
(11)
Где К′s – субстратная константа, учитывающая сродство фермента ко второй молекуле субстрата. Анализ этого уравнения, как и уравнения Михаэлиса-Ментен, целесообразно проводить раздельно в области низких ([S] < Кs´) и высоких ([S] ˃ Кm ) концентрациях субстрата.
При низких значениях концентрации субстрата уравнение упрощается до классического уравнения Михаэлиса-Ментен (рис. 2.12а), при линеаризации которого можно определить Кm, Vmax, kcat (k+2 ).
![]()
В области высоких концентраций субстрата уравнение принимает вид:
![]() (12)
(12)
Линеаризация этого уравнения в координатах (1/v, [S]) позволяет определить Vmax и Кs´ (рис.2.12б).
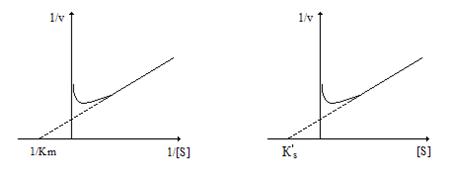
а) б)
Рис. 2.12. Линеаризация уравнения (12) в координатах (1/v 1/[S]) –а) и (1/v ; [S]) – б)
Зная величины Кm и Кs´, можно рассчитать оптимальную концентрацию субстрата по формуле:
![]() (13)
(13)
2.7. Задачи к разделу 2.
Задача 2.1. Исходя из данных табл. 2.1, определите характер ингибирования, Km и Vmax в присутствии и отсутствии ингибитора. Определить ингибиторную константу методом Диксона.
Таблица 2.1
[S], мМ | Скорость, мкмоль/мин | |||
Концентрация ингибитора | ||||
0 | 1,5 | 3,0 | 4,5 | |
0,20 | 0,32 | 0,17 | 0,12 | 0,09 |
0,73 | 1,10 | 0,60 | 0,35 | 0,35? |
1,26 | 1,50 | 0,87 | 0,77 | 0,60 |
1,78 | 1,96 | 1,20 | 0,81 | 0,65 |
2,31 | 2,22 | 1,61 | 1,12 | 0,98 |
2,84 | 2,40 | 1,62 | 1,40 | 1,31 |
3,36 | 2,85 | 2,00 | 1,51 | 1,35 |
3,90 | 2,65 | 1,80 | 1,73 | 1,44 |
4,42 | 2,80 | 2,56 | 2,12 | 1,47 |
4,94 | 3,24 | 2,58 | 1,82 | 1,72 |
Задача 2.2. Измеряли кинетику ферментативной реакции в зависимости от концентрации субстрата в присутствии или отсутствие ингибитора I и II. Были получены следующие данные (табл. 2.2):
Таблица 2.2
[S],M | Скорость, мкмоль/мин | ||
Без ингибитора | С ингибитором I | С ингибитором II | |
0,3 | 10,4 | 4,1 | 2,1 |
0,5 | 14,5 | 6,4 | 2,9 |
1,0 | 22,5 | 11,3 | 4,5 |
3,0 | 33,8 | 22,6 | 6,8 |
9,0 | 40,5 | 33,8 | 8,1 |
Задача 2.3. Определить Км и Vmax в присутствии и отсутствии ингибитора, исходя из данных, приведенных в табл. 2.3. Установить тип ингибирования.
Таблица 2.3
[S], мМ | v, ммоль/мин | |
контроль | ингибитор | |
1,25 | 1,72 | 0,98 |
1,67 | 2,04 | 1,17 |
2,50 | 2,63 | 1,47 |
5,00 | 3,33 | 1,96 |
10,00 | 4,12 | 2,38 |
Задача 2.4. Влияние оксамата натрия на скорость реакции, катализируемой лактатдегидрогеназой из Lactobacillis plantarum, изучали следующим образом.
2 мл трис-буфера, рН 8,0 и 0,2 мл 2 мМ раствора NADH добавляли в десять спектрофотометрических кювет. К пяти кюветам добавляли 0,1 мл 15 мМ раствора оксамата натрия. Во все кюветы добавляли различные объемы 0,6 мМ раствора пирувата натрия. Конечный объем проб доводили дист. водой до 2,9 мл. Реакцию запускали добавлением 0,1 мл раствора лактатдегидрогеназы измеряли оптическую плотность при 340 нм через мин после перемешивания и затем с интервалом в 1 мин. Полученные результаты оптических плотностей приведены в табл. 2.4.
Таблица 2.4
Время, мин | оптическая плотность при 340 нм | |||||||||
объем пирувата, мл | ||||||||||
без оксамата | с оксаматом | |||||||||
0,02 | 0,03 | 0,05 | 0,100 | 0,200 | 0,02 | 0,03 | 0,05 | 0,100 | 0,200 | |
1 | 0,800 | 0,804 | 0,776 | 0,772 | 0,726 | 0,788 | 0,803 | 0,803 | 0,780 | 0,760 |
2 | 0,776 | 0,768 | 0,721 | 0,633 | 0,583 | 0,776 | 0,784 | 0,776 | 0,729 | 0,683 |
3 | 0,750 | 0,727 | 0,668 | 0,544 | 0,460 | 0,764 | 0,765 | 0,748 | 0,681 | 0,608 |
4 | 0,724 | 0,692 | 0,617 | 0,460 | 0,350 | 0,751 | 0,745 | 0,721 | 0,627 | 0,532 |
Определить тип ингибирования. Рассчитайте величины КМ и VMAX в присутствии и без ингибитора.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 |
