
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Таблица 2.25
[S], 10-5 М | v/[E0], c-1 | [S], 10-5 М | v/[E0], c-1 |
1,98 | 4,15 | 20,0 | 7,50 |
2,50 | 4,65 | 50,0 | 6,91 |
3,00 | 5,13 | 100 | 5,72 |
4,00 | 5,89 | 200 | 4,22 |
6,00 | 6,63 | 300 | 3,30 |
8,00 | 7,15 | 396 | 2,74 |
10,0 | 7,15 |
Задача 2.26. Скорость реакции гидролиза )-(бензоилглицин) -2- оксиизовалериановой кислоты, катализируемой карбоксипептидазой А, уменьшается в присутствии избытка субстрата. Исходя из данных табл. 2.26, определите кинетические и равновесные параметры. Условия опыта: рН 7,5; 250 С; [Eo] = 6,1 10-10 – 1,2 10-8 М.
Таблица 2.26
[S] 10-4, M | v/[E]o, с-1 | [S] 10-4, M | v/[E]o, с-1 | [S] 10-4, M | v/[E]o, с-1 |
0,400 0,445 0,500 0,572 0,667 0,800 1,000 1,410 1,780 | 9,57 10,50 11,50 12,80 14,50 16,70 19,60 25,00 28,00 | 2,00 3,16 4,00 5,62 10,00 10,60 16,80 17,80 24,00 | 29,2 33,8 35,6 36,2 34,7 33,4 29,2 28,4 24,8 | 31,6 44,7 56,2 74,1 100,0 141,0 178,0 316,0 562,0 | 21,00 16,70 15,00 12,50 10,60 8,35 7,50 5,83 4,80 |
Задача 2.27. При изучении кинетики гидролиза монофенилфосфата, катализируемого щелочной фосфатазой, было найдено, что субстрат является одновременно и ингибитором ферментативной реакции. На основании данных табл. 2.27 определить кинетические и равновесные параметры ферментативного гидролиза. Условия опыта: Рн 10,0; 370 С; [E]o = 4 10-3 мг/мл.
Таблица 2.27
[S] 10-3, М | V 10-5 , моль мин-1 |
0,5 1,0 2,0 4,0 5,0 6,0 7,0 10 12 | 1,60 2,80 4,10 4,65 4,64 4,36 3,45 2,60 1,84 |
Задача 2.28. При увеличении концентрации субстрата в реакции гидролиза N-карбобензокси-L-триптофана, катализируемого карбоксипептидазой, скорость ферментативной реакции проходит через максимум, затем снижается и достигает постоянного уровня (табл.2.28). Определить значения кинетических и равновесных параметров реакции. Условия опыта: рН 7,5; 50 С; 0,04 М фосфатный буфер; 0,5 М KCl.
Таблица 2.28
[S]10-3 М | v/[E]o, c-1 | [S]10-3 М | v/[E]o, c-1 |
2,00 2,22 2,50 2,86 3,33 4,00 5,00 6,67 10,00 | 7,70 8,33 9,00 9,80 10,80 11,80 12,80 13,90 14,90 | 20,0 39,0 50,0 70,0 76,0 98,0 101,0 130,0 150,0 | 15,0 13,6 12,8 12,0 11,8 10,9 11,0 10,9 10,8 |
Раздел 3. Ферменты, не подчиняющиеся кинетике Михаэлиса-Ментен
Не все ферменты подчиняются кинетике Михаэлиса-Ментен. Для некоторых из этих ферментов кривая Михаэлиса имеет сигмоидную, а не гиперболическую форму. Обычно к этим ферментам относятся аллостерические ферменты, отличающиеся от обычных ферментов по ряду признаков:
1) имеют олигомерную структуру, т. е. обладают 4-й структурой и состоят из субъединиц, объединенных в единое целое связями слабого характера;
2) наряду с активными центрами имеют специальные регуляторные (или аллостерические) центры. Активные и аллостерические центры пространственно разобщены. К аллостерическим центрам могут присоединяться низкомолекулярные соединения, называемы эффекторами или модуляторами. В качестве эффекторов могут выступать промежуточные метаболиты или конечные продукты метаболических путей;
4) молекула эффектора, связываясь в аллостерическом центре, вызывает изменение конформации фермента. Это приводит в свою очередь к изменению конформации активного центра, в результате активность фермента увеличивается (активация) либо уменьшается (ингибирование). Эффекторы обратимо взаимодействуют с аллостерическими центрами. Они не обладают явным сходством с субстратами, а фермент проявляет исключительно высокую специфичность к молекуле эффектора;
5) у аллостерических ферментов наблюдается отклонение зависимости скорости ферментативной реакции (v) от концентрации субстрата (S) или аллостерического эффектора (Э), а именно отклонение от простых кинетических закономерностей типа гиперболического закона Михаэлиса-Ментен.
Термин аллостерический был предложен Ж. Мано и Ф. Жакобом в 1961 г. Они предположили, что взаимодействие эффектора с аллостерическим центром вызывает в молекуле фермента конформационные изменения, затрагивающие активный центр, в результате чего активность фермента изменяется.
Не все ферменты в клетке являются аллостерическими, но многие аллостерические ферменты являются ключевыми ферментами, определяющими скорость лимитирующих стадий обменных процессов в клетке.
Для аллостерических ферментов характерным свойством является наличие кооперативных эффектов: присоединение первой молекулы соответствующего лиганда (субстрата к активному центру или эффектора к аллостерическому центру) сопровождается конформационными изменениями, которые изменяют его сродство к субстрату или эффектору.
Кооперативные эффекты подразделяют на гомотропные и гетеротропные. Гомотропные эффекты, при которых взаимодействия с лигандами могут быть кооперативными и антикооперативными, наблюдаются для идентичных лигандов, например, для молекул субстрата (а также для молекул кофермента или ингибитора). Гетеротропные эффекты, при которых взаимодействия, также являющиеся либо кооперативными, либо антикооперативными, наблюдаются для молекул различных лигандов.
Кооперативные эффекты, кроме того, подразделяют на положительные и отрицательные (антикооперативные). В случае положительной кооперативности присоединение первой молекулы лиганда вызывает конформационные изменения в белковой молекулы, которые передаются и облегчают присоединение последующих молекул лиганда (при этом зависимость скорости реакции от концентрации лиганда является S-образной) (рис. 3.1).

Рис. 3.1. Зависимость скорости реакции от концентрации субстрата
При антикооперативности присоединение первой молекулы лиганда вызывает такие конформационные изменения в белке, которые затрудняют присоединение последующих молекул лиганда к соответствующим центрам (зависимость скорости реакции от концентрации лиганда является гиперболической, но это не равнобочная гипербола Михаэлиса-Ментен) (рис. 2.5.1).
Такие различия легко увидеть, если линеаризовать зависимости скорости реакции от концентрации лигандов для трех случаев: положительной и отрицательной кооперативности и кинетики Михаэлиса-Ментен (рис. 3.2).
При построении графика в координатах двойных обратных величин S-образная кривая преобразуется в кривую выпуклую к оси абсцисс. В последние годы было показано, что отдельные ферменты обнаруживают гиперболическую зависимость скорости от концентрации субстрата. Однако форма гиперболы в этих случаях отличается от классической гиперболы Михаэлиса-Ментен: в координатах Лайнуивера-Берка последняя преобразуется не в прямую линию, а в кривую выпуклую к оси ординат

Рис. 3.2. Линеаризация кривых, приведенных на рис. 3.1
У аллостерических ферментов, так же как и у нерегуляторных ферментов, наблюдается «насыщение» субстратом (рис. 3.3).
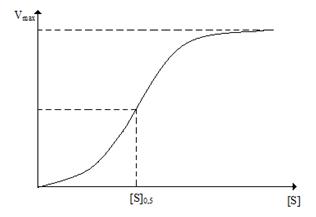
Рис. 3.3. Кривая с насыщением для аллостерического фермента
Хотя на сигмоидной кривой насыщения субстратом для аллостерических ферментов можно найти точку, в которой скорость реакции равна половине от максимальной скорости, эта величина не соответствует величине Km, поскольку поведение аллостерических ферментов не описывается гиперболической зависимостью, вытекающей и уравнения Михаэлиса-Ментен.
Первым белком с олигомерной структурой, проявляющим кооперативные эффекты при связывании кислорода, был гемоглобин. Кооперативность у гемоглобина была установлена Бором задолго до обнаружения подобных свойств у какого-либо фермента. Выявлению кооперативных свойств гемоглобина при его функционировании помогло сравнение с миоглобином, белком, не проявляющим кооперативных свойств и служащим для запасания кислорода в мышцах.
Впервые кооперативные эффекты были описаны Хиллом при анализе кривой насыщения гемоглобина кислородом, им же было предложено уравнение для описания этих кооперативных взаимодействий.
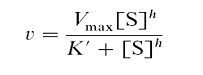
Где, v – наблюдаемая скорость ферментативной реакции;
VMAX – максимальная скорость реакции при концентрации субстрата, стремящейся к бесконечности;
К′– концентрация субстрата, при которой скорость равна половине от максимальной. Поскольку К′ не соответствует величине Км в данном случае вместо символа Км используют символы ([S]0,5, К0,5);
h – суммарное число связывающих центров в олигомерной молекуле фермента или коэффициент Хилла, или коэффициент кооперативных взаимодействий. Является мерой силы кооперативных взаимодействий при связывании лиганда. Коэффициент Хилла должен быть целым числом, но экспериментально определяемые величины часто не являются таковыми. В этих случаях за число связывающих участков принимают следующее, большее по величине целое число.
Например, экспериментально определенная величина h составляет 1,65. Это говорит о наличии минимум двух связывающих сайтов и что сайты проявляют уровень кооперативности выше среднего. Однако нельзя сказать, что фермент имеет только два связывающих центра. У него может быть три или 4 или более связывающих центров со слабо выраженной кооперативностью. Поэтому величина два в этом примере говорит о минимальном числе возможных связывающих центров.
3.1 Методы определения коэффициента Хилла
Существуют два метода определения коэффициента Хилла: метод, предложенный самим Хиллом и метод, разработанный с соавторами. В первом случае уравнение Хилла логарифмируется и преобразуется, приобретая следующий вид
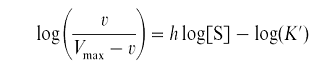
Это уравнение используют для построения графика, из которого можно определить величину h. График строится в координатах lоgv/(VMAX – v) против lg[S].
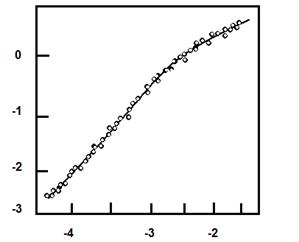
Пример линеаризованного графика Хилла приведен на рис. 3.4. Степень сигмоидности на линейных участках позволяет выявить силу кооперативности между центрами в олигомерном ферменте.
|
|

Рис. 3.4. Линеаризованная сигмоидная зависимость
Определение коэффициента Хилла по логарифмической зависимости требует знания Vmax. Однако не всегда эта величина известна.
с соавт. предложили так называемый разностный метод определения коэффициента Хилла. В этом случае строится график в координатах 1/v от log [S] (рис. 3.5).
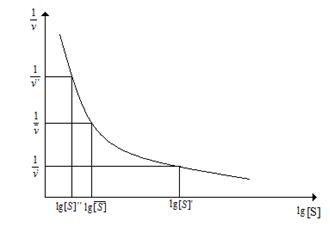
Рис. 3.5. Определение коэффициента Хилла «разностным» методом
Для нахождения коэффициента Хилла на кривой 1/v от log [S] выбираются три точки с координатами {1/v′ ; log [S]′}, {1/v; log [S]} и {1/v′′; log [S]′′}. Значения [S] для крайних точек отличаются отличаются от значений [S] для средней точки на постоянный множитель, обозначаемый ϰ (ϰ> 1): [S]′= [S]/ϰ и [S]′′=[S]ϰ.
Коэффициент Хилла рассчитывают далее по формуле:
h= log{(1/v′ - 1/v)/ (1/v - 1/v′′)}/ logχ
Суть этих преобразований сводится к следующему:
Разности обратных величин уже не содержа параметра Vmax, а отношение разностей при определенном выборе значений ![]() ; [S]ʹ; [S]ʹʹ - не содержит как параметра Vmax и [S]0,5.
; [S]ʹ; [S]ʹʹ - не содержит как параметра Vmax и [S]0,5.
Поэтому этот способ определения коэффициента Хилла получил название разносного метода.
После нахождения параметра коэффициента Хилла, остальные два параметра Vmax и [S]0,5 могут быть определены путем представления экспериментальных данных в координатах ![]() в соответствии со следующим линейным соотношением (рис. 3.6).
в соответствии со следующим линейным соотношением (рис. 3.6).
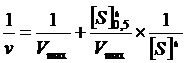
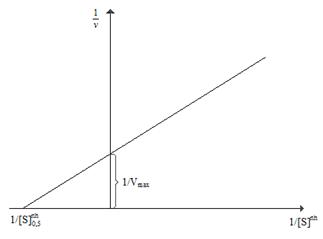
Рис. 3.6. Линеаризация уравнения Хилла с учетом величины коэффициента Хилла в координатах Лайнуивера-Берка
Коэффициент Хилла является параметром уравнения Хилла и не имеет физического смысла. Однако при достаточно сильных взаимодействиях коэффициент Хилла близок n (число активных центров в молекуле фермента). Таким образом, h ≤ n.
3.2. Определение коэффициента крутизны Кошланда
Оценить кооперативные взаимодействия в молекуле аллостерического фермента возможно с помощью коэффициента крутизны – Rх, предложенного Кошландом.
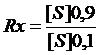 ,
,
где [S]0,9 – концентрация субстрата, при которой достигается 0,9 от VMAX; [S]0,1 – концентрация субстрата, при которой достигается 0,1 от VMAX.
Концентрации субстратов, при которых достигается 0,1 и 0,9 от максимальной скорости реакции можно определить, построив график зависимости скорости реакции от концентрации субстрата. Найдя по графику максимальную скорость, рассчитываем 0,1VMAX и 0,9VMAX и, опустив перпендикуляр на ось абсцисс, находим величины [S]0,1 и [S]0,9.
Для ферментов, подчиняющихся кинетике Михаэлиса-Ментен, коэффициент крутизны равен 81. Ферменты, характеризующиеся положительной кооперативностью, имеют коэффициент крутизны меньше 81, а отрицательной кооперативностью – больше 81.
Между коэффициентом Хилла и коэффициентом крутизны существует определенная зависимость:
Rx = 811/h,
В случае, когда h=1, уравнение Хилла превращается в уравнение Михаэлиса-Ментен.
Отношение [S]0,9/[S]0,1, индекс кооперативности, является обратной мерой кооперативных взаимодействий. Другими словами, большая разность в концентрации субстрата требуется для перехода от v=0,1Vmax до v=0,9Vmax, большие величины [S]0,9/[S]0,1 свидетельствуют о слабой степени кооперативности между центрами.
Так как коэффициент Хилла и индекс кооперативности (коэффициент крутизны) для олигомерных ферментов связаны друг с другом, вместе они позволяют измерить степень кооперативности между связывающими центрами и минимальное число этих взаимодействий. В случае гомотропных взаимодействий для молекул субстрата график зависимости v от [S] является сигмоидной кривой.
Это отражает ситуацию, когда с ферментом связываются несколько молекул субстрата, при этом связывание первой молекулы некоторым образом ускоряет связывание второй молекулы, т. е. при связывании с ферментом более чем одной молекулы субстрата наблюдается кооперативный эффект.
Такое поведение системы предполагает, что существует пороговая концентрация субстрата, ниже которой изменение его концентрации оказывает относительно небольшое влияние на скорость ферментативной реакции, однако при превышении порога даже небольшие изменения концентрации субстрата оказывают значительное влияние. Следовательно, в пределах относительно узкой области концентрации субстрата скорость ферментативной реакции изменяется очень резко, что весьма важно для аллостерических ферментов, поскольку многие из них являются регуляторными.
Для доказательства аллостерической природы положительной кооперативности по субстрату можно использовать прием десенсибилизации фермента. Для этого можно осуществить кратковременное его нагревание. Например, десенсибилизацию аспартаткарбамоилтрансферазы Герхард и Парди осуществляли в течение 4 мин при 60◦С, а UDP-глюкозо-4-эпимераза в течение 2 мин при 41◦С. Десенсибилизация может быть вызвана другими физическими воздействиями или химическим модифицированием фермента. Шанжё одним из первых добился десенсибилизации треониндегидратазы из E.coli по отношению к аллостерическому ингибитору – L-изолейцину при помощи n-хлормеркурийбензоата. Для десенсибилизации аллостерических ферментов (фосфофруктокиназы, глутаматдегидрогеназы и аспартаткиназы) осуществляли облучением растворов ферментов рентгеновскими лучами. После десенсибилизации ферменты не теряли каталитическую активность, но утрачивали возможность регулироваться аллостерическими эффекторами.
3.3. Задачи к разделу 3.
Задача 3.1. Аспартат-карбамоилтрансфераза катализирует следующую реакцию:
Аспартат + карбамоилфосфат → карбамоиласпартат + ортофосфат
и является первым ферментом при синтезе СТР. Активность фермента определяли по освобождению неорганического фосфата через определенные промежутки времени. Данные опытов, в которых определялась начальная скорость реакции при различных концентрациях аспартата и фиксированной концентрации карбамоилфосфата (3 мМ), приведены в табл. 3.1.
Таблица 3.1
Концентрация аспартата, мМ | Скорость, мкмольфосфата/ч | |
нативный фермент | фермент после прогревания при 600 в течение 4 мин | |
1,0 | 0,15 | 0,4 |
2,0 | 0,25 | 0,6 |
2,5 | 0,3 | 0,85 |
4,0 | 0,7 | 1,2 |
5,0 | 1,1 | 1,4 |
7,5 | 1,7 | 1,95 |
10,0 | 2,2 | 2,4 |
15,0 | 2,95 | 3,0 |
20,0 | 3,05 | 3,1 |
Определить кинетические параметры: коэффициент Хилла, коэффициент крутизны, [S]0,5, Vmax для нативного и прогретого фермента.
Задача 3.2. Аспартат-карбамоилтрансфераза катализирует следующую реакцию:
Аспартат + карбамоилфосфат → карбамоиласпартат + ортофосфат
и является первым ферментом при синтезе СТР. Активность фермента определяли по освобождению неорганического фосфата через определенные промежутки времени. Данные опытов, в которых определялась начальная скорость реакции при различных концентрациях аспартата, фиксированной концентрации карбамоилфосфата (3 мМ) и фиксированной концентрации СТР (0,1 мМ) приведены в табл. 3.2.
Таблица 3.2
Концентрация аспартата, мМ | Скорость, мкмольфосфата/ч | |
нативный фермент | Нативный фермент + 0,1 мМ СТР | |
1,0 | 0,15 | 0,025 |
2,0 | 0,25 | 0,10 |
2,5 | 0,3 | 0,125 |
4,0 | 0,7 | 0,35 |
5,0 | 1,1 | 0,5 |
7,5 | 1,7 | 1,03 |
10,0 | 2,2 | 1,6 |
15,0 | 2,95 | 2,5 |
20,0 | 3,05 | 2,9 |
Определить кинетические параметры для нативного фермента и фермента в присутствии СТР: коэффициент Хилла, коэффициент крутизны, Vmax, [S]0,5.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 |
