

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Экспрессия генов | Названия праймеров | Последовательность (5′ - 3′) |
β-Actin | β-act-f β-act-r | TGG AAT CCT GTG GCA TCC ATG AAA C TAA AAC GCA GCT CAG TAA CAG TCC G |
TLR2 | tlr2m-f tlr2m-r | CAG CTT AAA GGG CGG GTC AGA G TGG AGA CGC CAG CTC TGG CTC A |
TLR4 | tlr4m-f tlr4m-r | AGT GGG TCA AGG AAC AGA AGC A CTT TAC CAG CTC ATT TCT CAC C |
IL-4 | il4-f il4-r | CGA AGA ACA CCA CAG AGA GTG AGC T GAC TCA TTC ATG GTG CAGCTT ATC G |
IFN-γ | ifnγ-f ifnγ-r | AGC GGC TGA CTG AAC TCA GAT TGT AG GTC ACA GTT TTC AGC TGT ATA GGG |
ПЦР проводили на амплификаторе «МС-2 Терцик» (ДНК-технология, РФ). Для анализа продуктов амплификации использовали электрофорез в 2% агарозном геле с маркером молекулярных масс по 100 п. н. «PCR marker with Loading Dye» (Amresco, США). Продукты амплификации визуализировали при помощи системы «Gel Doc» («BioRad», США). Учет результатов ПЦР-анализа проводился по наличию или отсутствию на электрофореграмме специфических полос амплифицированной ДНК.
Иммуносерологические методы. Для определения концентрации суммарного иммуноглобулина Е (IgE) в сыворотке крови, цитокинов IL-4 и IFN-γ в сыворотке крови и супернатантах клеток применяли твердофазный иммуноферментный анализ с использованием коммерческих тест-систем производства (г. Санкт-Петербург) и -БЕСТ» (п. Кольцово). Оптическую плотность измеряли при длине волны 450 нм на микропланшетном фотометре для иммуноферментного анализа StatFax-3200 (Awareness Technology, США).
Иммунологические методы. Перитонеальные макрофаги, клетки селезенки, тимуса биомоделей получали общепринятым методом [РД ]. Мононуклеарные клетки выделяли из гепаринизированной крови путем центрифугирования в градиенте плотности 1,077 г/мл с применением Lymphocyte Separation Medium (Flow Laboratories, Великобритания). Жизнеспособность выделенных клеток оценивали с применением красителя трипанового синего. Реакцию лейкоцитолиза проводили в соответствии с МУ 3.1.2007-05 «Эпидемиологический надзор за туляремией». В качестве антигена использовали коммерческий препарат «Тулярин». Оценку внутриклеточного кислородзависимого метаболизма перитонеальных макрофагов осуществляли методом спонтанного НСТ-теста [, , 2001].
Определение апоптотических и пролиферирующих клеток проводили методом проточной цитофлуориметрии. Окраску ДНК иммунокомпетентных клеток митрамицином (Serva, Германия; Sigma, США), бромидом этидия (Serva, Германия) осуществляли по методу B. Barlogie et al. (1976). Измерения выполняли на импульсном проточном цитофлуориметре ICP-22 PHYWE (Германия), оснащенном 2048-канальным амплитудным анализатором импульсов Ortho Instruments (USA) модели 2102 для автоматической сортировки клеток и построения гистограмм.
Статистические методы. Статистическую обработку полученных данных проводили с использованием пакета программ Microsoft Excel. Определяли среднее значение (М), стандартную ошибку (m). Достоверность различий оценивали с использованием t-критерия Стьюдента. Результаты считали достоверными при значении р ≤ 0,05.
результаты исследований и их обсуждение
Для объективной интерпретации полученных результатов по действию изучаемых штаммов и антигенов на экспрессию TLR2 и TLR4 на первом этапе исследований была проведена оценка иммунобиологической активности (токсичности, иммуногенности, протективности) препаратов капсульного антигена чумного микроба, выделенных из рекомбинантного штамма Y. pestis КМ 277, вакцинного штамма Y. pestis EV НИИЭГ, а также протективного антигенного комплекса (ПАК) туляремийного микроба различных подвидов.
Согласно полученным данным все указанные препараты защищают экспериментальных животных (аутбредных мышей) от гибели в условиях моделирования чумной и туляремийной инфекции, не оказывают в дозе 100 мкг, используемой на последующих этапах исследования, токсического действия и не вызывают по данным проточно-цитофлуориметрического мониторинга повреждения иммунокомпетентных клеток по типу апоптоза.
Установлено, что протективная активность капсульного антигена (Ф1), полученного из рекомбинантного штамма Y. pestis КМ 277, в 3 раза выше (p< 0,05) по сравнению с препаратами капсульного антигена, выделенного из вакцинного штамма Y. pestis EV НИИЭГ (Ф1 серия 32 и Ф1 серия 44).
Значения ImD50 (ImD50 min ÷ ImD50max) для Ф1 из штамма Y. pestis КМ 277 и Ф1 серии 32, Ф1 серии 44 составили 8,3 (3,8÷19,0) мкг и 29,0 (26,2÷34,3) мкг, 28,94 (20,1÷32,5) мкг соответственно.
В ходе дальнейших исследований установлено, что введение штаммов
Y. pestis с различной генетической характеристикой, вакцинного штамма F. tularensis 15 НИИЭГ голарктического подвида, а также антигенов чумного и туляремийного микробов (капсульного антигена из рекомбинантного штамма Y. pestis КМ 277, вакцинного штамма Y. pestis EV НИИЭГ, протективного антигенного комплекса туляремийного микроба различных подвидов) оказывает влияние на уровень и кинетику экспрессии иРНК TLR2, TLR4 в клетках врожденного (перитонеальные макрофаги) и адаптивного (спленоциты) иммунитета биомоделей, степень выраженности которых зависит от вводимого антигена и вида возбудителя, из которого он был выделен.
Подкожное введение рекомбинантного штамма Y. pestis КМ 277 (ЕV11МpFSK3) Kmr – продуцента капсульного антигена чумного микроба Ф1, препарата Ф1 из штамма Y. pestis КМ 277, а также экспериментальной химической чумной вакцины индуцирует уже в первые часы повышение экспрессии иРНК TLR2 в спленоцитах мышей линии BALB/c (рисунок 1) с последующим усилением пролиферативной активности в центральном (тимусе) и периферическом (селезенке) органах иммунной системы и формированием напряженного протективного иммунитета (рисунок 2).
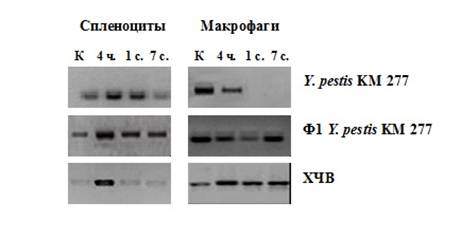
Рисунок 1 – Уровни иРНК TLR2 при иммунизации мышей BALB/c рекомбинантным штаммом Y. pestis KM 277, препаратом Ф1 Y. pestis KM 277 и химической чумной вакциной
Рисунок 2 – Относительное содержание (в %) пролиферирующих (>2 С ДНК) спленоцитов мышей линии BALB/с в условиях введения Ф1 Y. pestis KM 277, Ф1 серии 32, Ф1 серии 44
Примечание: * - р < 0,05
Уровень экспрессии TLR2, TLR4 в спленоцитах во все исследуемые сроки после иммунизации вакцинным штаммом Y. pestis EV НИИЭГ (Pgm-, pFra+, pCad+, pPst+), препаратами капсульного антигена, выделенного из агаровой культуры вакцинного штамма Y. pestis EV НИИЭГ (Ф1 серия 32), из бульонной культуры Y. pestis EV НИИЭГ (Ф1 серия 44) практически не отличался от показателей в контроле. В перитонеальных макрофагах биомоделей зарегистрировано кратковременное (через 4 ч) повышение экспрессии TLR4 в ответ на введение углеводсодержащего препарата Ф1 серии 32 в отличие от Ф1 серии 44, являющейся чистым белком, которое исчезало через 24 ч после иммунизации инбредных мышей.
В результате проведенного сравнительного анализа экспрессии TLR2, TLR4 и экспрессии генов цитокинов Th1(IFN-γ) и Th2 (IL-4)–зависимого ответа у биомоделей, индуцируемой вакцинным штаммом Y. pestis EV НИИЭГ, рекомбинантным штаммом Y. pestis КМ 277, химической чумной вакциной, капсульным антигеном чумного микроба, выделенным из Y. pestis КМ 277 и Y. pestis EV НИИЭГ, вакцинным штаммом F. tularensis 15 НИИЭГ, ПАК туляремийного микроба различных подвидов получены новые данные, свидетельствующие о преимущественной экспрессии клетками врожденного и адаптивного иммунитета иРНК TLR2 и IFN-γ в условиях in vivo.
Экспрессия иРНК IL-4 спленоцитами и перитонеальными макрофагами инбредных мышей, иммунизированных штаммами чумного микроба Y. pestis KM 277, Y. pestis EV НИИЭГ, препаратами капсульного антигена, выделенного из указанных штаммов, была ниже детектируемого уровня в течение всего периода наблюдения, что свидетельствует о превалировании Th1-зависимого ответа в процессе формирования специфической резистентности к чуме.
Введение экспериментальным биомоделям препарата протективного антигенного комплекса из вакцинного штамма F. tularensis 15 НИИЭГ голарктического подвида уже через 4 ч вызывало повышение экспрессии спленоцитами как TLR2, так и TLR4. Однако препараты протективных антигенных комплексов, полученные из вирулентного штамма голарктического (F. tularensis 503/840) и среднеазиатского (F. tularensis А179) подвидов, индуцировали усиление экспрессии только TLR2 в сочетании с IFN-g. Препарат протективного антигенного комплекса из штамма F. tularensis В399 A'Cole неарктического подвида не вызывал изменений экспрессии исследуемых типов Toll-подобных рецепторов, однако индуцировал на ранние сроки усиление экспрессии IFN-g и IL-4.
Важным результатом работы является проведенное впервые сравнительное изучение влияния вакцинации против чумы и туляремии на экспрессию иРНК TLR2, TLR4 и продукцию цитокинов клетками врожденного и адаптивного иммунитета у людей и оценка возможности применения полученных данных для разработки критериев оценки уровня противочумного иммунитета у людей.
Полученные в ходе выполнения работы данные о конститутивной экспрессии у людей гранулоцитами и мононуклеарными клетками крови иРНК TLR2 и TLR4 полностью согласуются с результатами других исследователей [Hornung V. и др., 2002; Zarember K.A., Godowski P. J., 2002], определивших с применением ПЦР-анализа в режиме реального времени уровни экспрессии различными субпопуляциями клеток периферической крови людей иРНК десяти Toll-подобных рецепторов, первые позиции среди которых по количеству синтезируемой иРНК заняли TLR2 и TLR4.
Впервые показано, что вакцинация людей против чумы и туляремии отечественными живыми вакцинами вызывает неоднозначные изменения уровня экспрессии иРНК TLR2 и TLR4 в лейкоцитах периферической крови.
Если в гранулоцитах через 3 месяца после вакцинации на фоне сформировавшегося адаптивного иммунитета транскрипционная активность генов TLR2 и TLR4 не была существенно изменена, то в мононуклеарных клетках введение живой чумной вакцины, но не живой туляремийной вакцины, приводило к снижению до недетектируемого уровня экспрессии иРНК TLR4, не влияя на уровень синтезируемой иРНК TLR2. Сочетанная вакцинация индуцировала экспрессию иРНК TLR2 и TLR4 как гранулоцитами, так и мононуклеарными клетками вакцинированных доноров. Зарегистрировано появление в результате одновременного введения живой чумной и живой туляремийной вакцин выраженной экспрессии иРНК TLR2 и TLR4 гранулоцитами у волонтера с недетектируемым до вакцинации уровнем экспрессии иРНК данных TLRs (рисунок 3).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
