
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

КОЭНЗИМ Q10 И ИДЕБЕНОН В ЛЕЧЕНИИ БОЛЕЗНЕЙ ДЫХАТЕЛЬНОЙ ЦЕПИ: ОБОСНОВАНИЕ И СРАВНИТЕЛЬНЫЙ АНАЛИЗ ЭФФЕКТИВНОСТИ
Резюме
Терапия пациентов с заболеваниями, обусловленными недостаточностью дыхательной цепи митохондрий, все еще остается по сути поддерживающей, несмотря на значительные успехи в идентификации молекулярных основ и понимании патогенеза митохондриальных болезней. Хиноны являются единственными фармакологическими агентами, применение которых дает определенный положительный эффект, но это заявление справедливо только для случаев, связанных с довольно специфическими дефектами дыхательной цепи. В данной статье коротко представлены данные о структуре, происхождении и распределении коэнзима Q10 в организме человека, далее суммируются все современные знания о его некоторых физиологических функциях. Затем обсуждается целесообразность применения различных видов хинонов для лечения митохондриальных болезней. Наконец, коротко оцениваются данные, полученные при лечении митохондриальных болезней с помощью хинонов, которые выступают в качестве заменителей переносчиков электронов или антиоксидантов.
1. Коэнзим Q10
Тканевые уровни коэнзима Q10 (CoQ10; убихинон 50) варьируют в широких пределах. В организме человека концентрации CoQ10 в сердце, почках, печени (114, 66,5 и 55 мкг/г ткани, соответственно) и в тонком кишечнике, толстом кишечнике, яичках и легких (11,5, 10,7, 10,5 и 7,9 мкг/г ткани, соответственно) различаются на порядок [1]. В скелетной мускулатуре, поджелудочной железе, щитовидной железе, селезенке и головном мозге концентрации коэнзима Q10 находятся на уровне средних значений (44, 33, 24,7, 24,6 и 13,4 мкг/г ткани, соответственно). Есть сообщения об общей тенденции к снижению уровня коэнзима Q10 в тканях по мере старения организма, однако возможны варианты в зависимости от типа исследуемой ткани [1]. При физиологических условиях ключевым параметром является редокс-потенциал CoQ10, а не его абсолютное количество [2]. Теоретически снижение концентрации CoQ10 (или, возможно, повышение окислительного потенциала) может отражать снижение уровня окислительного метаболизма в тканях из-за уменьшения потребности в транспорте электронов и активности антиоксидантной системы, особенно в связи с возрастными изменениями организма. Или, согласно более модной гипотезе, такое снижение может быть одним из ключевых событий в каскаде процессов, которые в конечном итоге приводят к возрастному сокращению окислительных и антиоксидантных ресурсов организма. Другими словами, снижение уровня коэнзима Q10 можно рассматривать и как приспособление к уменьшению метаболических потребностей организма, и как прогрессирующий с возрастом недостаток. На основе последней гипотезы возник несколько иррациональный и явно преждевременный энтузиазм по поводу чудодейственного воздействия коэнзима Q10 на процессы старения. Несомненным в этом было только процветание фирм-дистрибьюторов, а реальную пользу от употребления CoQ10 здоровыми людьми надо еще доказать.
2. Роль убихинона в митохондриях
Коэнзим Q10 играет важную роль в дыхательной цепи митохондрий, распределяя электроны между различными дегидрогеназами и цитохромными сегментами в дыхательной цепи (Рис. 1А) [3]. Пул CoQ10 намного превосходит все остальные компоненты дыхательной цепи (ДЦ) и является кинетически разнородным, а его окислительно-восстановительный потенциал строго регулирует активность дегидрогеназ (Рис. 1В) [4]. Функционирование пула СоQ10 объясняет нелинейную зависимость подавления электронного транспорта в ДЦ от концентрации антимицина – специфического ингибитора активности комплекса III (Рис. 1В; вставка) [5]. Наконец, через так называемый цикл Q (см. Рис. 6) функционирование СоQ10 обеспечивает вытеснение протонов из матрикса в межмембранное пространство по мере продвижения электронов через комплексы дыхательной цепи [6].
В зависимости от редокс-потенциала и протонового статуса, СоQ10, помимо участия в обмене электронов и протонов в дыхательной цепи, может вступать в реакцию с молекулярным кислородом с образованием супероксидов (Рис. 2) [1]. Так, при торможении антимицином потока электронов через комплекс III (см. Рис. 1В) происходит полное восстановление цитохрома типа b (этому способствует окисление цитохрома с1) и увеличивается количество нестабильного убисемихинона и, как следствие, супероксидов [7]. Поддержанию высокого мембранного потенциала и высокой скорости восстановления хинона с последующей повышенной продукцией супероксидов способствует как подавление АТФ-азы (олигомицином), так и дефект гена (мутация NARP, T8993G в гене митохондриальной АТФ-азы 6) [8]. Кроме прооксидантного эффекта убисемихинона, полностью восстановленный СоQ10 (убихинол) может удалять множество видов свободных радикалов, в том числе супероксиды, таким образом оказывая антиоксидантное действие (Рис. 2). Однако, поскольку СоQ10 является очень гидрофобным веществом, его природными мишенями являются преимущественно гидрофобные радикалы, такие как липопероксид или токоферил. Таким образом, СоQ10 может действовать и как прооксидантная молекула, и как антиоксидант [9].

Рис. 1. Предполагаемая роль убихинона в распределении электронов в дыхательной цепи митохондрий. (А) Различные дегидрогеназы во внутренней мембране митохондрии конкурируют за восстановление пула убихинона в дыхательной цепи; (В) из-за различий в специфической локализации и окислительно-восстановительных свойствах разных дегидрогеназ скорости потока электронов от дегидрогеназы к цитохромным сегментам дыхательной цепи также различны. Это определяет различия между дегидрогеназами в способности восстанавливать различные домены пула убихинона, который затем функционирует как кинетически разнородный (с разными скоростями обмена электронов в пуле, от медленной vs до быстрой vf). Однако при прогрессирующем подавлении цитохромных сегментов ингибитором (например, антимицином, который является специфическим ингибитором комплекса III) пуловая функция убихинона обеспечивает перераспределение электронов в оставшиеся неингибированные цитохромные сегменты. Пул хинонов затем ведет себя как кинетически гомогенный, что объясняет сигмовидный характер подавления дыхательной цепи антимицином, как схематично показано на вставке.
Интересные результаты получены в экспериментах на культурах человеческих фибробластов, выращиваемых в присутствии глюкозы или галактозы. Согласно заданным условиям эксперимента, уровень СоQ10 снижен настолько, что поток электронов по дыхательной цепи уменьшается, но не прекращается. При этом не наблюдали ни усиления в клетках окислительного стресса (через индукцию супероксиддисмутазы или повышение перекисного окисления липидов), ни повышения их чувствительности к окислительному стрессу, а это показывает, что антиоксидантная функция СоQ10, во всяком случае, в этих клетках (и в заданных условиях эксперимента), не имеет большого значения [10].
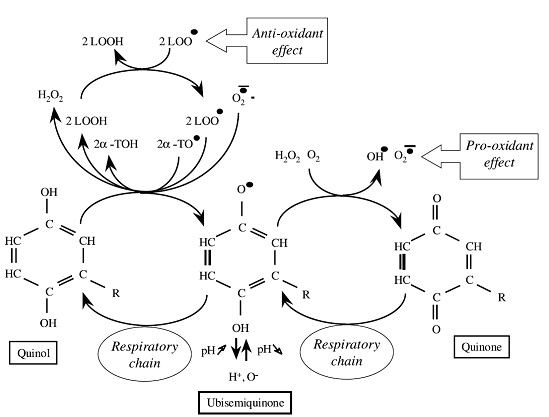
Рис. 2. Различные антиоксидантные и прооксидантные реакции, возможно, катализируемые хинонами. Прооксидантный эффект связан с образованием убисемихиноновых радикалов, которые легко вступают в реакцию с кислородом и его производными с образованием токсичных свободных радикалов кислорода. Антиоксидантная функция основана на образовании хинольной формы (полностью восстановленный хинон), которая может восстанавливать радикалы кислорода, липопероксид и токоферил, в зависимости от их локализации в мембранах. В мембране митохондрий окислительно-восстановительный потенциал хинонов контролируется активностью как цитохромных сегментов, так и нескольких дегидрогеназ, которые могут осуществлять восстановление пула хинонов, степень которого зависит от их локализации и окислительно-восстановительного потенциала.
Распределение молекул хинона по разным клеточным компартментам (во внутренней части или на поверхности мембран, в растворимых фазах) в значительной степени определяет их эффективность как антиоксидантов in vivo (Рис. 3). Манипуляции с гидрофобными свойствами хинонов привели к созданию большого количества его аналогов, нацеленных на разные мишени, что дает возможность оптимизации выбора препарата в зависимости от точной локализации радикалов, которые должны быть удалены из организма [11].
Наконец, получены прямые и косвенные данные, свидетельствующие о том, что СоQ10 вовлекается в некоторые дополнительные функции митохондрий, включая промежуточный метаболизм, пути биосинтеза и апоптоз, через регуляцию активности дегидрогеназ, генерацию супероксидов или прямое взаимодействие с другими компонентами митохондриальных мембран [12].
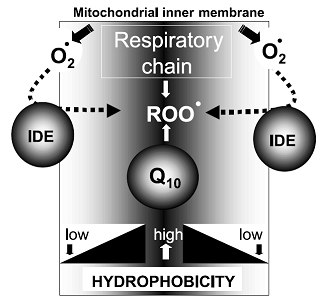
Рис. 3. Предполагаемая пространственная компартментализация антиоксидантного эффекта CoQ10 и идебенона в митохондриальных мембранах. В зависимости от структуры боковой цепи, распределение восстановленных хинонов в различных мембранных доменах (более или менее гидрофобных) определяет соответствующий сайт их антиоксидатной активности.
3. Экстрамитохондриальные функции убихинона
Внутриклеточный убихинон находится не только в митохондриях (приблизительно 1,4 мкг/мг белка; 1,86 мкг/мг во внутренних мембранах); например, самая высокая концентрация убихинона (2,62 мкг/мг белка) обнаруживается в пузырьках аппарата Гольджи клеток крысиной печени [13]. В этих клетках убихинон есть в лизосомах, плазматической мембране, пероксисомах и микросомах (1,86, 0,74, 0,29, 0,15 мкг/г белка, соответственно). Функция экстрамитохондриального убихинона пока точно не установлена. Предполагается, что окислительно-восстановительные циклы убихинона, ассоциированные со специфическими окислительно-восстановительными цепями мембран аппарата Гольджи и лизосом, способствуют одностороннему распределению протонов в этих органеллах [14]. В общем, убихинон в восстановленной форме, по-видимому, проявляет свою антиоксидантную активность во всех клеточных мембранах [1].
4. Экстрацеллюлярное действие убихинона
Помимо внутриклеточного действия, убихинон также может обеспечивать антиоксидантную защиту любого компонента, присутствующего в биологических жидкостях организма, который подвергается оксидативному повреждению. В частности, хиноны могут функционировать в крови, предотвращая (1) окисление липопротеинов низкой плотности (ЛНП), (2) свободнорадикальное повреждение при участии нейтрофилов при воспалительных заболеваниях и 3) оксидативное повреждение при участии эндотелиальных клеток, обусловленное ишемией-реперфузией [1]. В экспериментах на крысах и на людях получены убедительные доказательства того, что после перорального приема СоQ10, который слабо захватывается клетками, его концентрации в крови быстро нарастают [1]. Через экзогенные взаимодействия с поверхностью клеток циркулирующие в крови хиноны влияют на активность оксидоредуктазы плазматической мембраны (PMOR), которая катализирует восстановление цитозольным НАДН множества экстрацеллюлярных компонентов [15]. Так что вполне вероятно, что ожидаемая польза от применения СоQ10 действительно связана, хотя бы отчасти, с проявлением его активности в циркулирующей крови.
5. Биосинтез СоQ10
Биосинтез СоQ10 происходит путем конденсации парагидробензоатного кольца, получающегося из фенилаланина или тирозина, с 10-изопреноидной цепью (Рис. 4). Эта полипреноидная боковая цепь является производным мевалоната, который также является предшественником в биосинтезе холестерина [16]. Пренилированный парагидробензоат далее подвергается серии модификаций, включая реакции гидроксилирования, метилирования, О-метилирования и декарбоксилирования. Субклеточная локализация некоторых стадий этого процесса в организме человека пока неизвестна. На сегодняшний день описано всего несколько генов, имеющих отношение к этому процессу в организме человека, но пока врожденную недостаточность убихинона не связывают ни с одним из этих генов.


Рис. 4. Упрощенная схема биосинтеза коэнзима Q10. Точная внутриклеточная локализация некоторых стадий (цитозольная или митохондриальная) в организме человека остается неизвестной. Мевалонат используется большей частью как предшественник холестерина. Наконец, известна только часть генов, кодирующих ферменты, участвующие в этом биосинтезе в организме человека. Названия ферментов отмечены курсивом.
6. Коэнзимы Q
Молекула СоQ10 очень гидрофобна, это один из немногих антиоксидантов, которые могут синтезироваться в организме человека и после утилизации быстро регенерировать в восстановленную активную форму. В организме человека преобладает СоQ10, тогда как у крыс Rattus norvegicus [17] и у подсолнечника Helianthus tuberosus [18] обычно обнаруживается СоQ9. В различных живых организмах можно найти разные типы убихинона меньшей длины (СоQ6 в Saccharomyces cerevisiae) или большей (СоQ11 в растениях, относящихся к виду Capsicum) [16, 18]. Считается, что длина боковых цепей СоQ влияет на продолжительность жизни вида, но эта гипотеза пока не подтверждена экспериментально [19].
Помимо природных коэнзимов Q, существует целый ряд производных, которые были получены для повышения их биодоступности, точного нацеливания на конкретные клеточные отсеки и в надежде сохранить или даже улучшить предполагаемый положительный эффект от применения СоQ10 ([11] и ссылки). Однако единственными доступными фармакологическими препаратами пока остаются сам СоQ10 и идебенон − аналог СоQ10 с короткой цепью, отличающийся сниженной гидрофобностью.
7. Идебенон
Идебенон (6-(10-гидроксидецил)2,3-диметокси-5-метил-1,4-бензохинон; Рис. 5) представляет собой синтетическое вещество, первоначально запатентованное фирмой Takeda Chemical Industries (Осака, Япония). Разнообразные свойства этого соединения всесторонне проанализированы в обзоре Nagy в 1990 г. [20]. Лекарство появилось на японском рынке в 1986 г. Сначала основным показанием к применению идебенона было возрастное нарушение функций мозга. Идебенон обладает окислительно-восстановительными свойствами аналога хинона с возможным прооксидантным и антиоксидантным действием и способен взаимодействовать с другими носителями окислительно-восстановительных свойств, включая сам СоQ10, в дыхательной цепи митохондрий. ЛД50 идебенона для грызунов при внутрибрюшинном введении составляет примерно 10 г/кг веса тела, а при подкожном и пероральном введении ЛД50 повышается [21]. Обратимую подострую токсичность у грызунов наблюдали при введении дозы 2,5 г/кг/день [22], а доза 500 мг/кг день вызывала диарею у собак (в экспериментах использовали собак породы бигль) [24]. В исследованиях на хроническую токсичность, также проведенных на собаках, были получены сходные показатели, причем первые признаки токсичности появлялись при дозах 500 мг/кг веса тела, вводимых per os [23]. У идебенона не было обнаружено ни иммуногенных свойств на грызунах, ни мутагенного потенциала на бактериях и мышах. В организме человека идебенон быстро всасывается, концентрации в кровотоке повышаются в зависимости от введенной дозы (2 мкМ через
2 ч после перорального приема 100 мг), но также быстро снижаются за счет экскреции [24].
Безопасность идебенона для человека (60-300 мг) была установлена в долгосрочных испытаниях (2 года) на людях преклонного возраста, у которых не было зарегистрировано никаких значительных побочных эффектов [25]. Однако идебенон, как и другие аналоги хинона, является редокс-активным веществом с характерными кинетическими свойствами в отношении прочих носителей окислительно-восстановительного потенциала, включая компоненты комплексов дыхательной цепи. In vitro он способен конкурировать за электроны с природным СоQ10 в изолированных митохондриях и перенаправлять некоторую часть (величина варьирует) электронов за счет потока электронов к кислороду, особенно из комплекса I [26]. Быстро восстанавливаясь таким образом, идебенон оказывает мощное антиоксидантое действие на поверхности внутренних мембран митохондрий.
8. Обоснование применения хинонов для лечения митохондриальных болезней
Исходя из наших представлений о роли хинонов в митохондриях, следует различать попытки, направленные на (1) восстановление потока электронов в дыхательной цепи, (2) обеспечение электронами дыхательной цепи и (3) повышение митохондриальной антиоксидантной защиты.
8.1. Восстановление потока электронов
Для восстановления потока электронов необходимо, чтобы CoQ10, или его аналог, (1) достигал ткани-мишени, клетки и, в конечном итоге, митохондрий; (2) включался во внутреннюю мембрану митохондрий; (3) восстанавливал поток электронов путем "заполнения пробелов" (замещая пропущенные компоненты) или "создания мостиков" (в случае дефектности компонента) между носителями электронов. Фактически пока не доказана способность восстанавливать поток электронов при недостаточности дыхательной цепи in vitro или in vivo ни для CoQ10, ни для одного из его аналогов, за исключением случая дефицита CoQ10. В частности, обеспечение экзогенным CoQ10 или любым из аналогов с короткой цепью не может восстановить поток электронов in vitro через дефектный комплекс III. Примечательно, что гомологи с очень короткой цепью, такие как Q1 или Q0 (менадион), легко уводят электроны от дегидрогеназ дыхательной цепи. Но так как эти виды крайне нестабильны в восстановленном состоянии [27], электроны могут высвобождаться большей частью как свободные радикалы в растворимой фазе и, по-видимому, немногие из электронов могут повторно использоваться в цитохромных сегментах ДЦ.
8.2. Обеспечение дополнительным восстановительным потенциалом
Можно предположить, что в ряде конкретных случаев (дефицит комплекса I, II или III ДЦ) целесообразно обеспечить поступление в дефектную дыхательную цепь электронов из экзогенно восстановленных хинонов. In vitro экзогенно восстановленные хиноны легко окисляются изолированными митохондриями или пермеабилизированными клетками. Однако интактные клетки не могут захватывать восстановленные хиноны с достаточной для поддержания хоть сколько-нибудь значительного потока электронов скоростью [28]. Поэтому ценность такого подхода, по всей видимости, ограничена. Поскольку между аскорбатом и дыхательной цепью может циркулировать ограниченное количество попавшего в клетку хинона, проблему количественного захвата можно преодолеть путем добавления восстановителя (например, аскорбата) [29].

Рис. 5. Структурные особенности убихинона (CoQ10) и идебенона. При изображении молекулы CoQ10 в развернутом виде она кажется слишком длинной, чтобы соответствовать гидрофобному домену фосфолипидного слоя. Следует обратить внимание на короткую боковую цепь идебенона (10 атомов углерода, а в молекуле CoQ10 50 атомов углерода), насыщение боковой цепи идебенона, наличие терминальной карбоксильной группы и диметоксильных и метиловых остатков в кольце бензохинона. Все это делает идебенон менее гидрофобным, чем CoQ10, что влияет на его распределение и диффузию в биологических мембранах.
8.3.Укрепление системы антиоксидантной защиты
Применение хинонов представляется оправданным при наличии признаков образования свободных радикалов. Однако следует иметь в виду, что, поскольку свободные радикалы могут уводить электроны из дыхательной цепи и выполнять роль прооксидантов, обеспечение клеток хиноном не всегда безопасно. При определенных условиях человеческие клетки, культивируемые в присутствии экзогенных хинонов, могут быстро погибнуть. Лучше всего доставлять хиноны в клетку в окисленном состоянии, тогда они будут выполнять в митохондриях роль антиоксидантов, восстанавливаясь дыхательной цепью в хинол там (или максимально близко), где и происходит образование свободных радикалов (Рис. 3). Затем, во избежание автоокисления, семихинон, который образуется по мере восстановления свободных радикалов, должен пройти переокисление в дыхательной цепи (Рис. 6). Наконец, точное нацеливание хинонов на очаг образования свободных радикалов во внутренней мембране митохондрий определяет их фактическую эффективность. Оптимизация целевого воздействия хинонов на специфические мишени является объектом внимания активно развивающейся и очень перспективной области научных исследований [11].
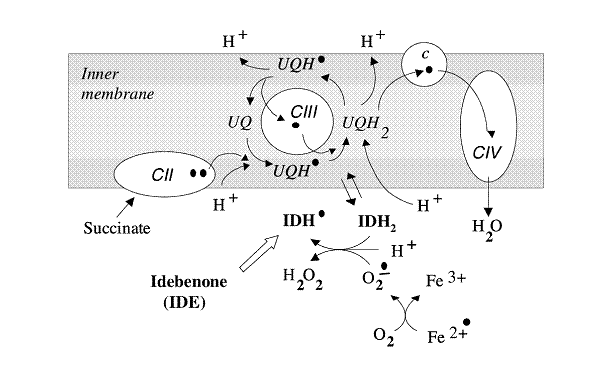
Рис. 6. Механизм антиоксидантного действия идебенона в митохондриях. После добавления к митохондриям идебенон восстанавливается дыхательной цепью, обеспечивая антиоксидантную защиту митохондриальных мембран от периферического оксидативного повреждения. Следует отметить, что в случае полной инактивации дыхательной цепи идебенон не будет оказывать никакого протективного действия.
Установленная на большом количестве испытуемых (пациенты с недостаточностью дыхательной цепи и здоровые испытуемые) безопасность длительного применения самого CoQ10 может быть связана с тем обстоятельством, что он просто не захватывается хоть в сколько-нибудь значительном количестве клетками с нормальным содержанием CoQ10. Из этого следует, что введение CoQ10 может быть нецелесообразно при внутриклеточном окислительном стрессе, причиной которого могла бы быть специфическая дисфункция дыхательной цепи. Этим можно объяснить и тот факт, что ни одно из проведенных двойных-слепых или открытых испытаний не продемонстрировало значительной эффективности CoQ10 для лечения заболеваний, обусловленных недостаточностью дыхательной цепи. Как говорилось выше, во многих случаях эффект CoQ10 на циркулирующие в крови маркеры окислительного стресса может быть обусловлен его прямым – экстрацеллюлярны – действием в биологических жидкостях организма.
При экспериментальном лечении специфических заболеваний дыхательной цепи используются оба качества хинонов, то есть его свойства носителей электронов и антиоксидантые свойства.
9. Заместительная терапия при истощении CoQ10
Сообщалось о снижении уровня CoQ10 при самых разных заболеваниях. Но считать доказанным, что эти изменения являются причиной или даже ключевым фактором в патогенезе этих заболеваний, преждевременно. Например, более или менее выраженное истощение запасов CoQ10 наблюдается при болезни Паркинсона [30], миотонической дистрофии [31], пародонтозе [32], бета-талассемии [33] и т. д. Также нет ясности в вопросе о том, действительно ли при снижении содержания CoQ10 меняется биоэнергетическая активность тканей или резервы антиоксидатной системы. Однако в литературе описано несколько доказанных случаев генетически обусловленного дефицита CoQ10, хотя молекулярную основу этих заболеваний выяснить пока не удалось.
В недавно опубликованном обзоре анализируются немногочисленные случаи предположительно первичного истощения CoQ10 в мышечной (и нервной) ткани [34]. При лечении CoQ10 клиническое состояние этих пациентов улучшалось, но в различной степени [34]. Позднее положительный эффект от применения CoQ10 описывался при некоторых случаях мозжечковой атаксии и мозжечковой атрофии, развитие которых объясняют истощением CoQ10 [35].
Только в одном случае у членов одной семьи был обнаружен значительный дефицит CoQ10 вследствие блокады биосинтеза, которая приводила к накоплению промежуточного продукта – пренилпирофосфата [28]. Двух больных детей успешно пролечили пероральным препаратом CoQ10. Сейчас (спустя четыре года) эти дети, у которых была серьезная энцефалопатия, а один из них был прикован к инвалидному креслу, могут самостоятельно ходить, бегать и совершать многокилометровые путешествия на велосипеде. Впечатляющий эффект от лечения коэнзимом Q10, достигнутый в данном конкретном случае, должен заставить понять, что необходимо делать все возможное для выявления первичных дефектов CoQ10, пока они не привели к развитию необратимых повреждений. С функциональной точки зрения, истощение CoQ10 лучше всего определяется в пробах путем измерения активности ферментов дыхательной цепи, которая в значительной степени зависит от функционирования пула CoQ10. Такими ферментами являются, например, глицерол 3-фосфат - и дигидрооротат-цитохром с-редуктаза [28]. Сукцинат-цитохром с-редуктаза в значительно меньшей степени зависит от пула CoQ10, хотя при его значительном истощении активность этого фермента также снижается. Добавление экзогенного аналога хинона (т. е. идебенона, децилубихинона, СоQ2) в тестируемые пробы восстанавливает нормальный уровень ферментативной активности. Следует отметить, что снижение зависимой от пула CoQ10 ферментативной активности также может быть связано с неправильным обращением с замороженными биологическими пробами. Поэтому для убедительного доказательства истощения CoQ10 рекомендуется исследовать свежий материал, а затем подтверждать результат, исследуя экстрагированные пробы количественным методом.
10. Наследственная атаксия Фридрейха
Применение хинона также открывает перспективы лечения угрожающей жизни кардиомиопатии, которая часто развивается при наследственной атаксии Фридрейха [36]. Атаксия Фридрейха является самой распространенной формой наследственной атаксии, возникает при экспансии ГАА-повтора в первом интроне гена, кодирующего фратаксин [37] – митохондриальный белок, который вероятнее всего участвует в синтезе или поддержании железо-серных центров ферментов [38-40]. В культивируемых человеческих фибробластах кожи снижение концентрации фратаксина затрудняет передачу сигнала супероксиддисмутазы [41]. О наличии окислительного стресса в митохондриях у пациентов, страдающих этим заболеванием, свидетельствует отсутствие активности митохондриальных ферментов, содержащих железо-серный центр [38], уменьшение ресурсов антиоксидантной системы [42] и увеличение доли окисленной ДНК в моче [43]. Так как известно, что идебенон (1) обладает мощными антиоксидантными свойствами, (2) проходит через гематоэнцефалический барьер и (3) легко захватывается клетками даже при нормальном уровне CoQ10, именно этот препарат, а не CoQ10, давали более чем 100 пациентам с наследственной атаксией Фридрейха, которые наблюдались в нашей клинике. Примечательно, что при исследовании активности дыхательной цепи в образцах различных тканей, взятых у этих пациентов, невозможно было обнаружить признаки истощения CoQ10. Тридцать восемь из этих пациентов получали препарат в течение более шести месяцев, значительное уменьшение размеров сердца (>20%) наблюдали примерно у 50% пациентов [44, 45]. За этот период ни у одного из получавших лечение пациентов не было выявлено значительного увеличения размеров сердца. Поскольку естественное течение заболевания неизбежно приводит к постепенному нарастанию гипертрофии сердца, которая может привести к дилатации сердца, эти результаты вполне обнадеживают в плане перспектив лечения этого заболевания. Также сообщалось об уменьшении доли окисленной ДНК в моче пациентов с атаксией Фридрейха, получавших идебенон [43]. Пока нет данных, на основании которых можно было бы сделать вывод об улучшении неврологического состояния больных при длительном сроке применения идебенона. Но, учитывая положительный эффект от применения препарата при кардиомиопатии и отсутствие побочных эффектов, следует рассмотреть возможность назначения препарата пациентам на стадии до появления клинической симптоматики. Поскольку известно, что (1) гипертрофия кардиомиоцитов может быть результатом выработки супероксидов и (2) атаксия Фридрейха является одним из немногих заболеваний, для которых повышенная продукция супероксидов подтверждена экспериментально, мы считаем, что в основе эффекта идебенона действительно лежит его выраженная способность удалять из организма супероксиды. Эффективность идебенона в плане профилактики митохондриальной кардиомиопатии при атаксии Фридрейха стала стимулом к тому, чтобы оценить эффективность его применения при митохондриальных миопатиях другой этиологии, возможно, связанной с дисфункцией пула хинонов (например, при снижении активности цитохром с-редуктазы). Описан удивительный случай улучшения состояния сердца одного пациента, принимавшего идебенон перорально; впоследствии этот пациент был исключен из списка кандидатов на пересадку сердца [46].
11. Окислительный стресс, митохондрии и хиноны
Повышенное образование супероксидов в митохондриях было продемонстрировано при целом ряде других форм недостаточности дыхательной цепи [8, 47]. Например, дефицит АТФ-азы, обусловленный мутацией NARP, связан с повышенной продукцией супероксидов, запускающей каскад апоптоза в культурах фибробластов кожи. Из экспериментов in vitro следует, что защитить клетки от апоптоза могут спиновые ловушки [8]. К сожалению, при дефиците АТФ-азы блокируется вся дыхательная цепь, так что применение редокс-цикличных антиоксидантов может дать только ограниченный эффект, поскольку для их функционирования необходим окислительно-восстановительный обмен с дыхательной цепью.
Апоптоз, запускаемый супероксидами, может также участвовать в довольно специфических (и не совсем понятных) процессах дегенерации зрительного нерва при наследственной атрофии зрительного нерва Лебера (LHON). Это заболевание связано с мутацией в митохондриальной ДНК. Фактически, долгое время это объясняли высокими и специфическими энергетическими потребностями зрительного нерва [48]. Очевидно, что передача сигнала является энергоемким процессом и поэтому предсказуемо чувствительна к любому значительному ограничению поступления энергии. Однако в культурах фибробластов кожи с гомоплазмией LHON (более 99% мутантных молекул митохондриальной ДНК с G11778A в гене ND4) не было выявлено ни снижения окисления субстрата, ни сокращения продукции АТФ (неопубликованные данные). Кроме того, вполне резонно предположить, что энергия требуется для нормального функционирования и других органов. Так что нужно найти другое объяснение селективной дегенерации зрительного нерва при болезни Лебера, и здесь альтернативой может быть высокая чувствительность к свободным радикалам кислорода, запускающим процессы гибели клеток. Исходя из известных антиоксидантных свойств CoQ10 и идебенона, эти препараты, иногда в комбинации с витамином Е, применяют для лечения пациентов с LHON. Уже в 1992 г. идебенон был заявлен как препарат, дающий определенный эффект при лечении атрофии зрительных нервов Лебера [49]. Однако вопрос об эффективности препарата при этом заболевании остается открытым, поскольку, несмотря на некоторые сообщения о положительном эффекте хинонов, проведение клинических испытаний до сих пор не организовано.
Во множестве клинических ситуаций ключевым событием считают оксидативные повреждения в результате дисфункции митохондрий. Активно изучаются последствия ишемии/реперфузии, связанные с митохондриальной дисфункцией и/или на нее влияющие, предполагается, что важную роль в этих процессах играет окислительный стресс [50]. Сообщалось об успешном применении как CoQ10, так и идебенона для предотвращения эффекта реперфузии при ишемическом повреждении, что, по-видимому, связано с антиоксидантными свойствами этих препаратов [51-53]. Оксидативными повреждениями центральной нервной системы на фоне дисфункции митохондрий часто пытаются объяснить нейродегенеративные заболевания [54] и процессы физиологического старения [55], хотя часто такие предположения не имеют твердых экспериментальных доказательств. Для лечения этих состояний также рекомендуют использовать и коэнзим Q10, и идебенон [56, 57], что открывает огромный коммерческий рынок для этих препаратов в развитых странах. Сообщений о токсичности коэнзима Q10 и идебенона пока нет, но нет и строгих доказательств того, что их применение полезно. На фоне первоначального энтузиазма были выдвинуты предположения о многочисленных полезных эффектах CoQ10, но они остаются гипотетическими, и чтобы их доказать, необходимо провести долгосрочные, рандомизированные, двойные-слепые испытания.
12. Вывод
Снижение уровня CoQ10, при котором может сокращаться поток электронов по дыхательной цепи и/или снижаться антиоксидантная защита, уверенно рассматривается не только в контексте редко встречающихся генетических дефектов, при которых нарушается биосинтез хинонов в клетках, но и в связи с множеством возрастных заболеваний и старением как таковым. Это позволило создать солидный рынок препарата коэнзима Q10 в так называемых развитых странах. Фактически, количество убихинона провозглашается ключевым фактором, определяющим продолжительность жизни различных организмов, хотя это заявление редко подкрепляется убедительными доказательствами. Вместе с тем, "наполнение" организма коэнзимом Q10 не кажется таким эффективным средством продления жизни человека. Какой-либо положительный эффект от его использования еще надо доказать потребителю, но, тем не менее, этот препарат уже ежедневно принимают до 10% пожилого населения некоторых стран северной Европы!
Однако, пусть даже в нескольких частных случаях, CoQ10 (или его аналоги) является одним из очень немногих веществ, которые демонстрируют неопровержимую эффективность при лечении митохондриальных болезней, таких как наследственная атаксия Фридрейха. Поэтому важно при скрининге на недостаточность дыхательной цепи не пропускать те случаи, которые отвечают на лечение хинонами.



НАРИСУНОЧНЫЕ НАДПИСИ
Рис. 1.
Дигидрооротат
Сукцинат
Алифатический ацил-КоА
вм
матрикс
Цитохромный сегмент
Рис. 2.
Антиоксидантный эффект
Прооксидантный эффект
Хинол
Дыхательная цепь
Убисемихинон
Хинон
Рис. 3.
Внутренняя мембрана митохондрий
Дыхательная цепь
низкая
высокая
ГИДРОФОБНОСТЬ
Рис. 4.
ацетил КоА
ацетоацетил КоА
3-ОН-метилглутарил КоА
мевалонат
5Ф-мевалонат
5ПФ-мевалонат
3Ф-5ПФ-мевалонат
изопентенил-ПФ
диметилаллил-ПФ
тирозин
4-ОН-фенилпируват
4-ОН-фениллактат
4-ОН-циннамат
4-ОН-бензоат
полипренилтрансфераза
ФПФС
геранил-ПФ
фарнезил-ПФ
геранилгеранил-ПФ
пентапренил-ПФ
гексапренил-ПФ
гептапренил-ПФ
октапренил-ПФ
нонапренил-ПФ
декапренил-ПФ
декапринил-ОН-бензоат
3-декапринил-4ОН-бензоат
3,4-дигидрокси-5-полипринилбензоат
3-метокси-4-гидрокси-5-полипринилбензоат
2-полипринил-6-метоксифенол
2-полипринил-6-метокси-1,4-бензохинон
2-полипринил-3-метил-6-метокси-1,4-бензохинон
2-полипринил-1,3-метил-5-гидрокси-6-метокси-1,4-бензохинон
Коэнзим Q10
Рис. 5.
Коэнзим Q10
56 Å: развернутая молекула СоQ10
50 Å: изопреноидная боковая цепь с 50 атомами углерода
Идебенон
(<50 Å: гидрофобный домен двойного фосфолипидного слоя)
Рис. 6.
Внутренняя мембрана
Сукцинат
Идебенон
Болезни
- Болезнь (Большая Советская Энциклопедия)
- Схема истории болезни. Методические указания для студентов II-III курсов медицинских институтов
- Детские болезни: периоды детского возраста
- Внутренние болезни
- Реферат на тему: Болезни, передающиеся наследственным путем
- Болезни почек
- Болезни органов дыхания
- Болезни зубов
- Болезни носа, горла и уха
- Болезни, передаваемые укусами насекомых
Проекты по теме:








 Основные порталы (построено редакторами)
Основные порталы (построено редакторами)