
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Аденовирусы - двухцепочечные ДНК-вирусы, инфицирующие покоящиеся и делящиеся клетки. В отличие от ретровирусов аденовирусы не интегрируются в геном хозяина и их ДНК сохраняется в ядре, как эписомальный элемент после инфицирования клеток хозяина. В процессе деления клеток введенная генетическая информация элиминируется, что не позволяет использовать аденовирусные векторы для переносов генов с высокой митотической активностью. Общее для всех аденовирусных векторов преимущество состоит в легкости отчистки и высокой эффективности инфицирования различных типов клеток хозяина, делящихся и неделящихся. Эти векторы могут нести трансгенную кассету емкостью 30 т. н. К недостаткам аденовирусных векторов относятся кратковременная экспрессия трансгена в большинстве тканей (от нескольких дней до недели) и относительно высокая иммуногенность.
Особое место среди вирусных векторов занимают генетические конструкции на основе вируса простого герпеса. Этот вирус не встраивается в геном клетки, локализируется вне ядра в виде эписом и подобно аденовирусам, создает малый риск вставочного мутагенеза. Вирус герпеса способен инфицировать неделящиеся клетки и многие клетки, не инфицируемые ретровирусами. Этот вирус имеет большой объем генома, что затрудняет его использование при конструировании векторных систем. Наиболее часто векторы, созданные на основе вируса простого герпеса, используются для трансдукции в клетки нервной ткани.
Ретровирусы ( РНК-вирусы ) располагают геномом с обратной транскриптазой, которая транскрибирует вирусный генетический материал в двуспиральную промежуточную ДНК, инкорпорирующую в ДНК хозяина.
Для конструирования используют ретровирусы типа С ( онкоретровирусы – вирус лейкоза мышей Молони, MuLV и вирус лейкоза птиц, ALV) лентивирусы (вирус иммунодефицита человека HIV) и спумавирусы. Модификация генома клетки с помощью ретровируса стабильна и переносится во все дочерние клетки, происходящей от трансфицированной клетки. Объем генетической информации, переносимой с помощью ретровирусных векторов, невелик и составляет не более 10-12 т. п.н. Уникальное свойство ретровирусов – способность инкорпорировать гликопротеины оболочки различных вирусов в свою липидную оболочку. Это свойство ретровирусов используют, когда клетка-мишень не располагает рецепторной структурой для предназначенного к встраиванию вектора. Упаковка вектора в чужую вирусную оболочку позволяет расширить панель клеток-мишеней для вирусного вектора. Ретровирусные векторы менее имуногенны чем другие вирусные вектора.
В ряде работ показано, что наибольшей эффективностью при вирусной трансдукции отличились системы, полученные на основе лентивирусного генома. Распространение получили векторы, произведенные на основе вируса иммунодефицита человека (ВИЧ), так как они позволяют заражать широкий спектр клеток, стабильно интегрироваться в геном, что приводит к долговременной экспресссии.
С помощью гликопротеина, входящего в состав оболочки, вирусная частица взаимодействует с белками (например, CD4), присутствующими на поверхности клетки, после чего вирион попадает внутрь клетки. Далее вирус разрушается и при этом происходит высвобождение одноцепочечной РНК в комплексе с некоторыми белками. Для интеграции лентивируса в клетки хозяина РНК вируса должна подвергнуться обратной транскрипции. В результате этого сложного процесса образуется двуцепочечная ДНК, способная встроиться в ДНК клетки, что происходит с помощью вирусного белка интегразы (кодируемые белком pol).(рис. 9,10) С этого момента в клетке могут образовываться мРНК, кодирующие вирусные белки. Экспрессия вирусных генов контролируется промотор-энхансерным элементом, расположенных в длинных концевых повторах провируса. В цитоплазму попадают короткие вирусные транскрипты, которые используются для синтеза вирусных белков Tat и Rev, помогающих увеличить уровень экспрессии вирусных генов и экспортировать из ядра РНК, не претерпевшую сплайсинг. В результате в цитоплазме накапливается РНК двух видов: мРНК полноразмерная (может заново упаковываться в вирусную частицу) и мРНК претерпевшую одиночный сплайсинг. Необходимо отметить, что продукция вирусных частиц приводит к гибели клеток.
Общая стратегия создания вирусных систем заключается в минимализации количества содержащих в конструкции (плазмиде) вирусных генов, а также в физическом разделении цис - и транс - действующих факторов. Целью является создание вирусных частиц, дефектных по репликации
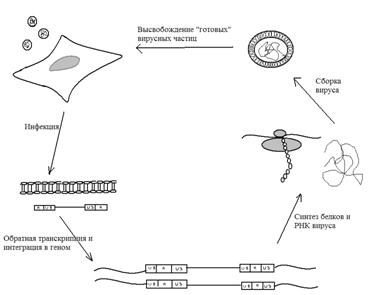
Рис. 9. Cхематическое изображение жизненного цикла лентивирусов. (Саложин, 2009)
Лентивирусные системы первого поколения создавались с использованием трех плазмид, которые кодировали все необходимые цис-элементы, все вирусные гены, кроме генов, кодирующих гликопротеин оболочки. Таким образом, формируется вирусная частица, имеющая все белки, необходимые для инфицирования клеток, но образующаяся РНК в результате транскрипции провируса кодирует только продукт вносимого гена.
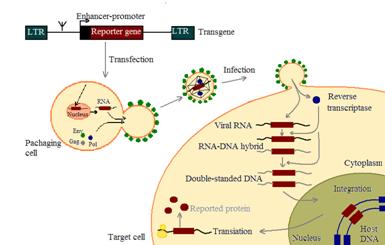
Рис 10 Размножение ретровирусного вектора в упаковывающих клетках и трансдукция в клетку-мишень. (Sikorski, 1998)
В лентивирусных системах второго поколения делеции подверглись дополнительные гены, необходимые для in vivo, но необязательные для культивирования in vitro.
В лентивирусных системах третьего поколения отсутствуют все вирусные гены, кроме тех, которые необходимы для образования вирусных частиц. Все эти изменения сделаны для того, чтобы вероятность образования способного к размножению вируса была бы близка к нулю. Также были сделаны различные дополнительные модификации, увеличивающие эффективность лентивирусных систем и безопасность работы с ними.
Для увеличения экспрессии трансгена используются различные стратегии: например элиминирование участков, которые потенциально могут подвергаться метилированию ДНК (что приводит к подавлению транскрипции); введение в вектор последовательностей, обеспечивающих высокую эффективность синтеза РНК трансгена: областей приклепления к ядерному матриксу, инсуляторов, тканеспецифичных промоторов.
Табл.2 Сравнение свойств вирусных векторов (Малайцев, 2009)
Показатели | Вектор | ||||
Аденовирус | Ретровирус | Лентивирус | ААВ | ВПГ | |
Размер частицы (нм) | 70 | 100 | 100 | 20 | 200 |
Тип и размер генома | дв. ДНК (36) | РНК (8,8) | РНК (9,6) | од. ДНК (4,7) | дв. ДНК (152) |
Интеграция в хромасомы | Нет | Да | да | да | нет |
Иммунный ответ | +++ | +/- | +/- | +/- | + |
Экспрессия | Кратковременная | Перманентная | Перманентная | Долговременная | От кратковременной до перманентной |
Возможные риски | Воспалительный ответ, цитотоксичность | Вставочный мутагенез, генерирование репликационно компетентных ретровирусов | Вставочный мутагенез, генерирование репликационно компетентных ретровирусов | Вставочный мутагенез | Воспалительный ответ, цитотоксичность |
Кроме значительно более высокой по сравнению с другими методиками трансдукция с использованием лентивирусных систем позволяет проводить более сфокусированные исследования и добиться большей специфичности воздействия, чем при экспериментах с фармакологическими агентами. Но необходимо помнить о возможном риске для исследователя.
Аденоассоциированный вирус не является патогенным, представляет собой одноцепочечную ДНК парвовируса с капсидом диаметром 26нм. Каждый конец одноцепочечной ДНК содержит инвертированный повтор, который является единственным цис-действующим элементом необходимым для репликации генома и упаковки. Геном несет два вирусных гена rep и cap. Вирус использует два промотора и альтернативный сплайсинг для кодирования четырех белков необходимых для репликации, третий промотор кодирует три белка вирусного капсида. При естественном инфицировании AAV необходим аденовирус или вирус простого герпеса для того, чтобы завершить свой жизненный цикл. Рекомбинантный AAV (rAAV) вектор доставки генов может быть получен путем удаления двух вирусных генов (rep и cap) и вставки трасген-кассеты между двумя инвертированными повторами. rAAV может успешно заражать и преобразовывать широкий спектр типов клеток, таких как нейроны, гепатоциты, миоциты и многие другие. Он способен инфицировать как покоящиеся, так и делящиеся клетки.
Глава 4. Использование маркерных генов при трансдукции и трансфекции
Трансдукция и трансформация могут применяться как для увеличения экспрессии белков, уже присутствующих в клетках, так и для снижения или полного подавления экспрессии определенных белков, а также для внесения конструкций, содержащих чужеродные гены или иные последовательности ДНК.
Для эффективного изучения трансфицированных клеток вектор должен содержать маркерный ген, позволяющий идентифицировать гибридные клетки или конкретные вещества в них. Можно выделить 2 группы маркерных генов, позволяющие отличить трансформированные клетки:
1. Селективные гены, отвечающие за устойчивость к антибиотикам Основной принцип работы такого маркера – способность трансформированных клеток расти на селективной питательной среде с добавкой определенных веществ, ингибирующих рост и деление нетрансформированных, нормальных клеток.
2. Репортерные гены, кодирующие нейтральные для клеток белки, наличие которых в тканях может быть легко протестировано. Чаще всего в качестве репортерных используются гены β-глюкуронидазы (GUS), зеленого флуоресцентного белка (green fluorescent protein - GFP), люциферазы (LUC), хлорамфениколацетилтрансферазы (CAT). К настоящему времени из этого арсенала наиболее часто используют гены GUS и GFP и, в меньшей степени, LUC и CAT.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
