
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Наибольшее распространение получил GFP (рис.11) из-за его особых свойств, а именно его способности флуоресцировать в видимой (зеленой) области спектра при облучении длинноволновым УФ. Эта флуоресценция обусловлена непосредственно белком, для ее проявления не требуется субстратов или кофакторов. Благодаря этому свойству ген GFP стал очень перспективным репортерным геном, позволяющим проводить разнообразные прижизненные (недеструктивные) исследования с трансгенными организмами и клетками. В 1960—1970-е года зелёный флуоресцентный белок был выделен Осаму Симомурой вместе с другим светящимся белком экворином из медузы Aequorea victoria. Оказалось, что в A. victoria взаимодействие ионов кальция с экворином вызывает голубое свечение белка. Часть этой биолюминесценции переносится на зелёный флуоресцентный белок, который поглощает синий свет и испускает флуоресценцию зелёного цвета, что приводит к сдвигу в свечении медузы в зеленую часть спектра. Однако применение GFP в молекулярной биологии началось лишь в 1990-х годах. В 1992 году Прэшер клонировал и секвенировал ДНК белка, после чего из-за недостатка финансирования вынужден был закрыть проект и разослал полученную ДНК в несколько лабораторий, в том числе в лабораторию Мартина Чалфи. Чалфи экспрессировал последовательность в Escherichia coli и Caenorhabditis elegans и опубликовал результаты в журнале Science в 1994. Месяц спустя были опубликованы независимые результаты из лаборатории Фредерика Тцуи. Оказалось, что GFP принимал нативную конформацию и образовывал флуорофор при комнатной температуре и без добавления дополнительных кофакторов, что обеспечило возможность использования белка в качестве маркёра в клетках многих организмов. Кристаллическая структура белка была расшифрована в 1996 году в лаборатории Ремингтона. Она прояснила механизм образования флуорофора и роль окружающих аминокислот. Это позволило получать мутантные GFP с повышенной устойчивостью, с различной флуоресценцией и другими улучшенными свойствами по сравнению с диким типом.
Сейчас создано огромное количество GFP-подобных белков с различными спектрами поглощения и излучения, что позволяет наблюдать сразу несколько клеточных клонов.

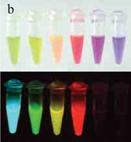
Рис.11 а – кристаллическая структура GFP; b – коллекция очищенных рекомбинантных белков при обычном освещении (вверху) и их флуоресценция (внизу) (Богданова и др., 2007)
Используя ген флуоресцентного белка (репортерный), находящийся под контролем какого-либо конкретного гена, можно изучать уровень экспрессии этого белка в живых клетках. Экспрессия подобных химерных белков (белков слияния) позволяет визуализировать трансфицированные клетки. Примером визуализации может служить так называемая «brainbow» мышь. При ее создании используется множество трансгенных кассет, кодирующих несколько флуоресцентных белков. Рекомбиназы случайным образом вырезают часть кодирующих их генов, в результате чего каждая клетка окрашивается своим цветом. Таким образом, если раньше мозг выглядел как однородная структура, то на такой трансгенной мыши можно изучать архитектуру нейронных связей, выбрав нейрон и проследив за его аксоном (рис.12).
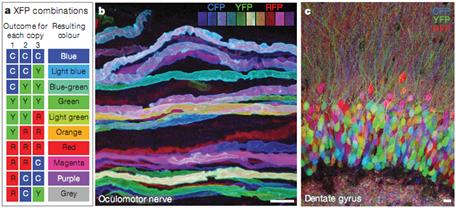
Рис. 12. а – экспрессия трех флуоресцентных белков при независимой рекомбинации может быть получено 10 различных цветов; b – различные области околомоторных аксонов; с – клетки зубчатой извилины (Livet et al, 2007)
Существуют так называемые флуоресцирующие таймеры, меняющие цвет флуоресценции по мере созревания белка. Это удивительное свойство позволяет отслеживать изменение экспрессии генов во времени. Использование белков слияния дает возможность исследовать движение флуоресцентных конструкций внутри клетки. Этот подход особенно актуален для исследования белков, меняющих свою локализацию в клетке при активации (инактивации) .
Еще одним примером использования трансдукции может служить изучение межбелковых взаимодействий с помощью FRET (Fluorescence resonance energy transfer, т. е. перенос энергии по индуктивно резонансному механизму), при котором происходит перенос энергии от донора к акцептору посредствам диполь-дипольного взаимодействия. С помощью FRET можно регистрировать любые биохимические сигналы, если две флуорофора находятся на определенном расстоянии друг от друга.
Возможна трансфекция конструкциями, содержащими искусственно созданные белки на основе GFP, люцеферазы или экворина, которые чувствительны к изменению различных внутриклеточных параметров, таких как определение уровня свободного цитоплазматического кальция, внутриклеточного АТФ, установление изменений концентрации водорода (рН), а также локализацию исследуемых конструкций в разных органеллах клетки, анализ активности различных биологически активных веществ, изучение сигнальных систем и рецепторов.
Используемые термины
Вектор - это общее название «транспортного средства» для целенаправленной доставки того или иного вещества, например гена.
Вставочный мутагенез – процесс неспецифического встраивания генетического элемента в геном, при котором могут активироваться онкогены, вследствие чего могут возникать опухоли.
Гибридизация ДНК, гибридизация нуклеиновых кислот — соединение in vitro комплементарных одноцепочечных нуклеиновых кислот в одну молекулу. При полной комплементарности объединение происходит легко и быстро, а в случае частичной некомплементарности слияние цепочек замедляется, что позволяет оценить степень комплементарности. Возможна гибридизация ДНК-ДНК и ДНК-РНК.
Лигирование — сшивание молекул ДНК посредством ферментов ДНК-лигаз.
Липосома – самопроизвольно образующийся липидный бислой, который
образует коллойдную частицу в водной среде
Отжиг (англ. annealing) — гибридизация фрагментов ДНК
Плазмиды или внехромосомный генетический элемент – кольцевая или линейная молекула нуклеиновой кислоты, расположенные вне хромосом и реплицирующая независимо от генома.
Полиплексы – комплексы ДНК и полимеров.
Трансдукция – перенос генетического материала из одной клетки в другую с помощью вируса.
Трансфекция - процесс введения нуклеиновой кислоты в клетки животных невирусным методом.
Трансформация – процесс введения нуклеиновой кислоты в бактериальные клетки невирусным методом.
Цис-элемент (цис-регуляторный элемент) — участок ДНК или РНК, который регулирует экспрессию генов, локализованных на той же молекуле ДНК (хромосоме). Цис-элементы обычно являются местами связывания одного или нескольких регуляторных белков (транс-факторами), кодируемых другими генами. Цис-элементы могут располагаться в 5'-концевом участке гена (в области промотора), в 3'-концевом участке (на нетранслируемых участках), а также в интронах.
Эписомы — генетические элементы бактерий, способные существовать как в интегрированном в бактериальные хромосомы состоянии, так и в виде автономных плазмид.
Эписомы – генетические элементы, способные существовать как в интегрированном в хромосомы состоянии, так и в виде автономных плазмид.
Список литературы
1. , , Киселев стволовых клеток и клеточные технологии //. Биология стволовых клеток и клеточные технологии: Учебная литература для студентов медицинских вузов. – М.: Медицина, Шико, 2009. том 1; 89-106 с.
2. Никитин введение веществ и органелл в клетку в технологиях клеточной инженерии // Цитология. - 2007. - Т.8. - С. 631-641.
3. , Большаков клеток нервной системы // Журнал высшей нервной деятельности. - 2009. - Т.1. - С. 3-14.
4. , Биохимия: В 3-х т. Т. 3. Пер. с англ.-М.: Мир, 1985
5. Barbu A., Welsh N. Lipofection of Insulin-Producing RINm5F Cells: Methodological Improvements // J. Liposome Res. - 2007. - V.17. - P. 49–62.
6. Felgner J. H., Kumar R., Sridhar C. N.,Wheeler C. J., Tsai Y. J., Border R., Ramsey P., Martin M., Felgner P. L. Enhanced gene delivery and mechanism studies with a novel series of cationic lipid formulations // J. Biol. Chem. - 1994. - Vol.4. - P.2550-2561.
7. Hu W. S., Pathak V. K. Design of retroviral vectors and helper cells for gene therapy//Pharmacol. Reviews. - 2000. - Vol.52. - P.493-512.
8. John A. O’Brien, Sarah C. R. Lummis. Encyclopedia of Medical Genomics and Proteomics. - 2006. - Vol.10. - P.1081
9. Lim M. L., Lum M. G., Hansen T. M., Roucou X., Nagley P. On the release of cytochrome c from mitochondria during cell death signaling // J. Biomed. Sci. - 2002. - Vol.6.Pt.1. - P.488-506.
10. Luo D.,Saltzman M. W. Synthetic DNA delivery systems // Nature Biotechnology.2000.Vol.18.P.33 – 37.
11. Patil S. D., Rhodes D. G., Burgess D. J. DNA-based therapeutics and DNA delivery systems: A comprehensive review // T. AAPS J. - 2005. - Vol.7. - P. E61-E77.
12. Plank C., Schillinger U., Scherer F., Bergemann C., Remy J. S., Krotz F., Anton M., Lausier J., Rosenecker J. The magnetofection method: using magnetic force to enhance gene delivery // Biol. Chem. - 2003. - Vol.5. - P.737-747.
13. Promega [Электронный ресурс]. Режим доступа.: http://www. /guides/transfxn_guide/transfxn. pdf
14. Shigekawa, K. and Dower, W. J. (1988) Electroporation of eukaryotes and prokaryotes: A general approach to the introduction of macromolecules into cells // BioTechniques. – 1988. – V.6, - P.742–51.
15. Sikorski R., Peters. R. Gene Therapy: Treating with HIV // Science. - 1998. – V.282 (5393) - P.1438a.
16. Teruel M. N., Blanpied T. A., Shen K., Augustine G. J., Meyer T. A versatile microporation technique for the transfection of cultured CNS neurons. // J. Neurosci. Methods. - 1999. – V.93 (1). – P. 37-48.
17. Tirlapur U. K., Konig K. Targeted transfection by femtosecond laser. Nature. - 2002. - V.418(6895). – P. 290-291.
18. , Общая и молекулярная генетика, Сибирское университетское издательство, Новосибирск: Сибирское университетское издательство, 2007. - 480 с.
19. Г97 Генетика / Бартон Гуттман, Энтони Гриффитс, Дэвид Сузуки, Тара Куллис. — Пер. с англ. О. Перфильева. — М.: ФАИР-ПРЕСС, 2004. - 448 с.
20. Гены; пер.9-го англ. изд – М.: БИНОМ. Лаборатория знаний, 2011. - 896 с.
Оглавление
Введение 3
Глава 1.Создание рекомбинантной ДНК 6
Глава 2. Трансфекция и трансформация 9
Глава 3. Трансдукция 16
Глава 4. Использование маркерных генов при трансдукции и трансфекции. 23
Используемые термины 26
Список литературы 27
МЕТОДЫ ГЕНЕТИЧЕСКОЙ ТРАНСФОРМАЦИИ
Составители:
Олеся Михайловна Широкова
Мария Валерьевна Ведунова
Учебное пособие
Государственное образовательное учреждение высшего
профессионального образования «Нижегородский государственный университет им. ».
603950, Нижний Новгород, пр. Гагарина, 23.
Подписано в печать. Формат 60х84 1/16.
Бумага офсетная. Печать офсетная. Гарнитура Таймс.
Усл. печ. л. 3,0. Уч.-изд. л. 3,3.
Заказ № Тираж 100 экз.
Отпечатано в типографии Нижегородского госуниверситета
им.
603600, 7
Лицензия ПД от 14.05.01

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
