
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
Нижегородский государственный университет им.
Национальный исследовательский университет
МЕТОДЫ ГЕНЕТИЧЕСКОЙ ТРАНСФОРМАЦИИ
Учебное пособие
Рекомендовано методической комиссией биологического факультета для студентов ННГУ, обучающихся по направлению 020400.62 «Биология», по профилю "Биофизика"
Нижний Новгород
2013
УДК 591.1.085.23 (075.8)
ББК Е991 я73
М54
М54 Методы генетической трансформации. Составители: , – Нижний Новгород: Издательство Нижегородского госуниверситета, 2013. 30 с.
Рецензенты: д. б.н., проф. ,
д. б.н. .
Учебное пособие содержит краткие теоретические сведения о некоторых современных методах генетической инженерии.
Руководство предназначено для студентов старших курсов биологического факультета в рамках курса «Нейрональные сети».
Ответственный за выпуск
председатель методической комиссии биологического факультета ННГУ, д. п.н., профессор
УДК 591.1.085.23 (075.8)
ББК Е991 я73
© Нижегородский государственный
университет им. , 2013
Введение
Открытия в области медицины и биологии 20-го века привели к снижению смертности и, как следствие, постоянному приросту населения планеты. Однако потребности постоянно растущего общества не могли быть удовлетворены традиционно применяемыми технологиями. Развитие молекулярной биологии и открытия в области генетики стали основой для постоянно продолжающегося роста и развития человеческой цивилизации. Генная инженерия не является наукой в широком смысле, но служит инструментом биотехнологии и предназначается для получения желаемых качеств у изменяемого или генетически модифицированного организма. Генная инженерия использует методы таких биологических наук, как молекулярная и клеточная биология, цитология, генетика, микробиология, вирусология. В отличие от традиционной селекции, в ходе которой генотип подвергается изменениям лишь косвенно, генная инженерия позволяет непосредственно вмешиваться в генетический аппарат. Примерами применения генной инженерии являются получение новых генетически модифицированных сортов зерновых культур, производство человеческого инсулина путём использования генномодифицированных бактерий, производство эритропоэтина в культуре клеток, создание новых экспериментальных животных для научных исследований.
Генетическая модификация отдельных клеток и целых организмов стала возможной благодаря мультидисциплинарным открытиям, сделанным во второй половине 20 века.
В 1944 году Эйвери, Мак Леод и Мак Карти показали, что носителем наследственной информации является ДНК. До этого момента считалось, что гены имеют белковую природу. С этого времени начинается интенсивное изучение нуклеиновых кислот. Спустя десятилетие, в 1953 году Уотсон и Крик создали двуспиральную модель ДНК. Именно этот год принято считать годом рождения молекулярной биологии.
На рубеже 50 - 60-х годов были выяснены свойства генетического кода, а к концу 60-х годов его универсальность подтверждена экспериментально. Разработаны методы выделения высокоочищенных препаратов неповрежденных молекул ДНК. ДНК вирусов и плазмиды вводили в клетки в биологически активной форме, обеспечивая их репликацию и экспрессию соответствующих генов. В 70-х годах открыт ряд ферментов (рестриктаз, ДНК-лигаз, транскриптаз), катализирующих реакции превращения ДНК.
Историю развития генетической инженерии можно условно разделить на три этапа.
Первый этап связан с доказательством принципиальной возможности получения рекомбинантных молекул ДНК in vitro. На этом этапе были получены гибриды между различными плазмидами; доказана возможность создания рекомбинантных молекул с использованием исходных молекул ДНК из различных видов и штаммов бактерий, их жизнеспособность, стабильность и функциональность.
Второй этап связан с началом работ по получению рекомбинантных молекул ДНК между хромосомными генами прокариот и различными плазмидами, доказательством их стабильности и жизнеспособности.
Третий этап - начало работ по включению в векторные молекулы ДНК (ДНК, используемые для переноса генов и способные встраиваться в генетический аппарат клетки-рецепиента) генов эукариот.
В настоящее время основным объектом биотехнологии является кишечная палочка (E. coli). Эта бактерия - поставщик огромного количества белков, необходимых человеку (гормоны, биологически активные белки, рекомбинантные белки иммунного ответа, кормовые белки и многие другие). Одним из первых белков, полученных из бактериальных клеток, был инсулин. Ранее инсулин получали из клеток поджелудочной железы животных, поэтому стоимость его была очень высока. Для получения 100 грамм кристаллического инсулина требуется 800-1000 килограмм поджелудочной железы, а одна железа коровы весит 200 - 250 грамм. Это делало инсулин дорогим и труднодоступным для широкого круга диабетиков. В 1978 году исследователи из компании "Генентек" впервые получили инсулин в специально сконструированном штамме кишечной палочки. Было показано, что выделенный и очищенный инсулин не содержит бактериальных белков, эндотоксинов и других примесей, не дает побочных эффектов (в отличии от инсулина животных), а по биологической активности от него не отличается. Из 1000 литров культуральной жидкости можно получать до 200 грамм гормона, что эквивалентно количеству инсулина, выделяемого из 1600 килограмм поджелудочной железы свиньи или коровы.
Генетическая инжене́рия (генная инженерия) — совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами и введения их в другие организмы. На данный момент генная инженерия широко применяется в экспериментальной биологии, в частности для трансфекции и трансдукции эукариотических клеток, или организмов, а так же для трансформации прокариотических клеток.
Трансфекция или трансдукция эмбриональных стволовых клеток (ЭСК) открывает огромные перспективы в изучении функций различных генов, направленной генетической модификации дифференцированных клеток, перепрограммирование их генома и возврат к плюрипотентному состоянию.
Для изучения функций генов, особенностей индивидуального развития, коррекции наследственных заболеваний удобно использовать такие экспериментальные модели, как трансгенные животные. В частности, внесения дополнительного генетического материала, манипуляции с вирусными векторами позволяют получать животных с пониженным уровнем экспрессии интересующих генов (нокдаун животные). Вирусные векторы также могут быть использованы для получения новых моделей различных нейродегенеративных заболеваний и генной терапии различных патологий.
Существуют различные методы внесения генетического материала в клетку. Вектор - это общее название «транспортного средства» для целенаправленной доставки того или иного вещества, например гена. Векторы бывают либо синтетическими (основаны на полимерных материалах, например липосомах), либо «натуральными», т. е. природного происхождения (например плазмиды или векторы на основе различных вирусов). В зависимости от выбранного вектора ген может быть интегрирован в геном клетки-мишени, локализован в ядре в качестве искусственной мини-хромосомы или, как эписомальный элемент, находиться в ядре или цитоплазме. Трансфекция - процесс введения нуклеиновой кислоты в клетки животных невирусным методом. Аналогичный процесс в бактериальных клетках называется трансформацией. Трансдукция – перенос генетического материала из одной клетки в другую с помощью вируса.
Глава 1.Создание рекомбинантной ДНК
Технология создания рекомбинантных ДНК – одно из важнейших достижений молекулярной биологии. В лабораторных условиях существует возможность создавать новые комбинации неродственных генов. Затем эти новые гены можно ввести в подходящие клетки и размножить (клонировать) с помощью механизмов синтеза ДНК клетки-хозяина. Основные этапы клонирования ДНК:
1. Создание рекомбинантной молекулы. Интересующий исследователя фрагмент ДНК ковалентно присоединяется к ДНК вектора. Основное свойство вектора состоит в том, что он может автономно реплицироваться в клетке хозяине. Молекулы ДНК можно соединить путем лигирования (лигирование - сшивание молекул ДНК посредством ферментов ДНК-лигаз).
2. Введение в клетку хозяина. Большинство бактериальных и эукариотических клеток поглощает «голые» молекулы ДНК из среды. Эффективность поглощения низка (примерно 1 на 106 молекул ДНК), но в специально подобранных экспериментальных условиях можно трансформировать значительную часть клеток. Существует и другой метод: клетки заражают реконструированными вирионами, содержащими рекомбинантные молекулы ДНК. В таком синтетическом вирусном геноме интересующий исследователя ген замещает участок вирусной ДНК, не имеющий существенного значения для репликации.
3. Отбор. Данный этап состоит в том, чтобы определить, какие клетки несут рекомбинантную молекулу ДНК, содержащую нужный ген. Такие клоны можно отобрать по признаку присутствия вектора или самого встроенного гена. Например, некоторые плазмидные векторы сообщают клетке устойчивость к какому-либо антибиотику. В качестве репортерных генов могут быть использованы гены, кодирующие флуоресцентные белки.
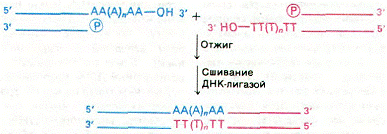
Молекулами ДНК можно легко оперировать in vitro с помощью рестриктирующих эндонуклеаз, ДНК-лигаз и других высокоспецифических ферментов, действующих на ДНК. При получении рекомбинантной ДНК вектор подготавливают к соединению с клонируемым фрагментом путем расщепления в определенном месте с помощью рестрикционной эндонуклеазы. Если с помощью этого фермента две цепи ДНК расщепить со смещением, то образуются комплементарные одноцепочечные концы (липкие концы). Второй метод соединения двух неродственных молекул ДНК основан на присоединении poly(dА)-фрагментов к 3'-концу одной молекулы и poly(dТ)-фрагмента 3'-концу другой молекулы (рис. 1). Такой метод получил название коннекторный метод.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
