
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Клонирование и экспрессия генов. Гены sps, spp, fruK и ams амплифицировали с геномной ДНК Mm. alcaliphilum 20Z и клонировали в векторах pET30a+, pET28b или pET22b+. Полученными рекомбинантными плазмидами pETsps, pETspp, pETfruK или pETams трансформировали компетентные клетки E. coli Rosetta (DE3). Экспрессию генов в E. coli индуцировали добавлением изопропил-1-тио-β-D-галактопиранозида (ИПТГ). Рекомбинантные ферменты SPS-His6, SPP-His6, FruK-His6 и Ams-His6 очищали металл-хелатной хроматографией на колонке с Ni2+-NTA-агарозой, согласно протоколу фирмы “Qiuagen” (Германия). Чистоту белковых препаратов и молекулярную массу субъединиц белков определяли электрофорезом в 8-12%-ном SDS-ПААГ по методу Лэммли [Laemmli, 1970]. Нативные молекулярные массы (м. м.) ферментов определяли с помощью градиентного 4-30% ПААГ электрофореза. Концентрацию белка определяли модифицированным методом Лоури [Schacterle, Pollack, 1973].
Характеристика сахарозофосфатфосфатазы. Активность SPP определяли по образованию неорганического фосфата двумя методами: непрерывно (с сопрягающими ферментами пирофосфат-зависимой 6-фосфофруктокиназой, фруктозофосфатизомеразой и глюкозо-6-фосфатдегидрогеназой) или периодически, используя реактив Бенцини [Bencini et al., 1983] при 35оС и рН 6,5. Для определения оптимума рН использовали следующие буферы: Na-цитратный (pH 5.0–6.0), MES-KOH (pH 6.0–7.0), Трис-HCl (pH 7.5–9.0) и Na-карбонатный (pH 9.0–10.0). Значения Km и Vmax рассчитывали с помощью программы SigmaPlot 10.
Характеристика амилосахаразы. Общую активность амилосахаразы определяли по скорости образования фруктозы, с помощью сопрягающих ферментов – фруктокиназы, фруктозофосфатизомеразы и глюкозо-6-фосфатдегидрогеназы. Гидролитическую активность измеряли по скорости образования глюкозы, используя сопрягающие ферменты – гексокиназу и глюкозофосфатдегидрогеназу. Трансгликозилирующую активность рассчитывали как разность общей и гидролитической активности. Измерения проводили при 30оС и рН 8,0. При необходимости в реакционную смесь добавляли гликоген в конечной концентрации 0,1 мг/мл. Для определения оптимума рН использовали следующие буферы: MES-KOH (pH 6.0–7.0), TAPS-KOH (pH 7.5–9.0) и Na-карбонатный (pH 9.0–10.0). Для определения термостабильности фермента аликвоты преинкубировали в микропробирках от 5 мин до 1 ч при 30, 40, 50 или 60°С, быстро переносили в лед, затем измеряли остаточную активность фермента при 30°С.
Характеристика фруктокиназы. Активность FruK определяли с помощью сопрягающих ферментов – фруктозофосфатизомеразы и глюкозо-6-фосфатдегидрогеназы. Влияние рН на активность FruK изучали с использованием следующих буферов (50 мМ): MES-KOH (pH 6,0–7,0), K-фосфатный (pH 6,5–5,5), Трис-HCl (pH 8,0–9,0) и Na-карбонатный (pH 9,0–10,5). Для определения термостабильности FruK аликвоту фермента преинкубировали от 5 мин до 1 ч при 30–60°С, быстро переносили в лед, затем измеряли остаточную активность фермента при 30°С.
За единицу активности ферментов принимали количество белка, катализирующее превращение 1 мкмоль субстрата в минуту.
Получение нокаут-мутантов. Фрагменты ДНК, содержащие нуклеотидные последовательности, фланкирующие гены sps, ams, ectB и ectC, клонировали в суицидальном векторе pCM184. В результате были получены векторы pCMsps, pCMams и pCMectBC. Для коньюгации в Mm. alcaliphilum 20Z использовали штамм-донор E. coli S17-1, который предварительно трансформировали соответствующей плазмидой. Трансконъюганты отбирали по устойчивости к канамицину и чувствительности к тетрациклину.
Выделение и анализ осмопротекторов. Низкомолекулярные органические осмопротекторы экстрагировали из клеток метанолом. Содержание сахарозы определяли антроновым методом [Handel, 1968]. Для количественного определения эктоина использовали модифицированный метод ВЭЖХ [Ешинимаев с соавт., 2007].
Все измерения были проведены в трех повторностях, на рисунках и в таблицах представлены средние значения и стандартные отклонения.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
1. Определение физиологической роли сахарозы у Mm. alcaliphilum 20Z.
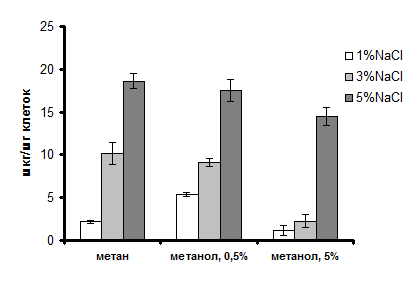 При росте Mm. alcaliphilum 20Z на метане или в присутствии метанола содержание сахарозы в клетках возрастало с увеличением солености среды (рис. 1). Выявленная зависимость накопления сахарозы от внешней солености удовлетворяет определению осмопротектора. Ранее было выдвинуто предположение, что процесс биосинтеза сахарозы у галофильных/толерантных метанотрофов может служить механизмом стока избыточного формальдегида, накапливающегося в среде в высоких концентрациях при экспоненциальном росте клеток на метаноле [Ешинимаев с соавт., 2002].
При росте Mm. alcaliphilum 20Z на метане или в присутствии метанола содержание сахарозы в клетках возрастало с увеличением солености среды (рис. 1). Выявленная зависимость накопления сахарозы от внешней солености удовлетворяет определению осмопротектора. Ранее было выдвинуто предположение, что процесс биосинтеза сахарозы у галофильных/толерантных метанотрофов может служить механизмом стока избыточного формальдегида, накапливающегося в среде в высоких концентрациях при экспоненциальном росте клеток на метаноле [Ешинимаев с соавт., 2002].
Рис. 1. Накопление сахарозы клетками Mm. alcaliphilum 20Z.
Однако, при замене метана на метанол (0,5% по объему) в среде культивирования, внутриклеточное содержание сахарозы у Mm. alcaliphilum 20Z не возрастало, а в клетках, выращенных в среде с низкой соленостью в присутствии 5% метанола, уровень дисахарида уменьшался (рис. 1).
Содержание сахарозы в клетках, растущих в присутствии 5% NaCl (0,855М), достигает 14-18 мкг/(мг биомассы), в пересчете на внутриклеточную воду это соответствует приблизительно 50 мМ, что недостаточно для уравновешивания осмотического давления среды. Вместе с тем известно, что при той же солености Mm. alcaliphilum 20Z накапливал эктоин в концентрации около 700 мМ [Khmelenina et al., 1999]. Логично предположить, что у данного метанотрофа сахароза является дополнительным к эктоину осмопротектором.
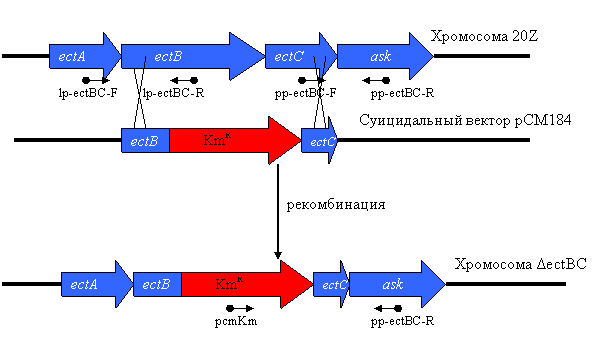
 Рис. 2. Схема инактивации генов ectB и ectC и расположение праймеров.
Рис. 2. Схема инактивации генов ectB и ectC и расположение праймеров.
Для подтверждения осмопротекторной функции сахарозы у Mm. alcaliphilum 20Z, с помощью двойной гомологичной рекомбинации нами был получен штамм ΔectBC с делецией по генам ectB и ectC, кодирующим, соответственно, ферменты биосинтеза эктоина – диаминобутиратаминотрансферазу и эктоинсинтазу (Рис. 2).
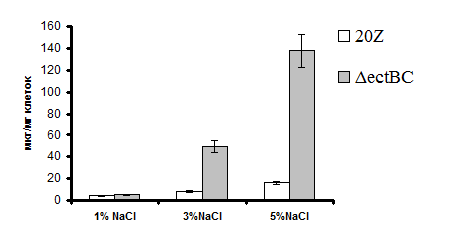
Рис. 3. Накопление сахарозы клетками штаммов 20Z и ΔectBC, выращенных при различных концентрациях NaCl.
В клетках штаммов 20Z и ΔectBC, выращенных в среде с метанолом (0,5% по объему) при различной солености, определяли содержание эктоина и сахарозы. Анализ показал, что мутантный штамм ΔectBC не накапливал эктоин, однако концентрация сахарозы в клетках мутанта была в ~8 раз выше, чем в клетках дикого типа (Рис. 3).
Ранее близкие результаты были получены для мутантного штамма Synechocystis sp. PCC 6803, дефектного по гену АДФ-глюкопирофосфорилазы и неспособного синтезировать основной осмопротектор – гликозилглицерин. Содержание сахарозы у мутанта было в 29 раз выше, по сравнению с диким типом, и соизмеримо с уровнем гликозилглицерина у дикого штамма [Miao et al., 2002]. Это указывает на присутствие у галотолерантных организмов компенсаторных механизмов, позволяющих восполнять недостаток одного осмопротектора за счет увеличения концентрации другого.
2. Идентификация и филогенетический анализ генов метаболизма сахарозы
В недавно опубликованном частично аннотированном геноме Mm. alcaliphilum 20Z (http://www. genoscope. cns. fr/externe/English/corps_anglais. html) обнаружен локус из 4 генов, продукты которых, предположительно, участвуют в метаболизме сахарозы. Транслированные аминокислотные последовательности этих генов проявляли гомологию с сахарозофосфатсинтазами (SPS), сахарозофосфатфосфатазами (SPP), сахарокиназами pfkB-семейства и амилосахаразами (Ams). Предполагаемая сахарозофосфатсинтаза Mm. alcaliphilum 20Z проявляла наибольшее сходство с ранее охарактеризованными ферментами из цианобактерий Synechocystis PCC 6803 [Lunn et al., 2003], Synechococcus sp. 7002 [Cumino et al., 2010] и Prochlorococcus marinus [Lutfiyya et al., 2007] (соответственно 56,8; 53,1 и 42% идентичности транслированных аминокислотных последовательностей). Значительно меньшее сходство обнаружено с сахарозофосфатсинтазами таких растений, как Nicotiana tabacum [Lutfiyya et al., 2007], Arabidopsis thaliana [Park et al., 2008] и Zea mays [Bruneau et al., 1991] (29; 28,9 и 28,4% идентичности), и SPS галотермофильной анаэробной бактерии Halothermothrix orenii (20,9% идентичности) [Huynh et al., 2005].
Продукт второго гена был гомологичен сахарозофосфатфосфатазе из Synechococcus sp. 7002 [Cumino et al., 2010] (49,8% идентичности), однако имел лишь незначительное сходство с SPP Zea mays и N. tabacum (15,9 и 16, 7 % идентичности) [Lunn et al., 2000; Chen et al., 2005].
Открытая рамка считывания (ОРС), следующая за предполагаемым геном сахарозофосфатфосфатазы, кодировала белок, сходный с сахарокиназами pfkB-семейства. Данное семейство включает АТР-зависимые ферменты: рибокиназу, кетогексокиназу, фруктокиназу, 2-дегидро-3-дезоксиглюкокиназу, 1-фосфофруктокиназу, тагатозокиназу, 6-фосфофруктокиназу архей, а также минорную 6-фосфофруктокиназу (PfkB) E. coli [Bork et al., 1993; Sigrell et al., 1997].
Продукт четвертой ОРС данного кластера был аннотирован в геноме изучаемого метанотрофа как α-амилаза, однако имел наиболее высокую гомологию с охарактеризованными амилосахаразами гетеротрофных бактерий: Alteromonas macleodii [Ha et al., 2009], Neisseria polysaccharea [De Montalk et al., 1999], Deinococcus geothermalis [Jung et al., 2009] и Deinococcus radiodurans [Pizzut-Serin et al., 2005] (39,4; 38,5; 37,6 и 36,7% идентичности соответственно).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
