
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Таблица 4. Свойства некоторых фруктокиназ прокариот.
Параметры | Mm. alcaliphilum 20Z | Thermococcus litoralis | Bifidobacterium longum | Zymomonas mobilis | Lactococcus lactis |
Молекулярная масса субъединицы, кДа | 35 | 35 | 35 | 28 | 32 |
Субъединичная структура | мономер | димер | мономер | димер | димер |
рН-Оптимум | 9,0 | 7,5-8,0 | 6,0 | 7,4 | 7,0 |
t-Оптимум | 60o | 80o | 50o | ||
Субстратная специфичность | Фруктоза | Фруктоза | Фруктоза | Фруктоза Манноза | Фруктоза Манноза |
Km, мМ: Фруктоза ATP | 0,25 1,3 | 2,3 0,81 | 0,74 0,77 | 0,7 0,45 | 0,31 0,59 |
Vmax, Е/мг белка: Фруктоза АТФ | 141 | 920 | 1,8 | 350 | - |
Относительная активность с донором фосфорильных групп | АТФ (100%), ГТФ (52%), УТФ (22%), ЦТФ (0%), ФФн(0%) | АТФ (100%), ИТФ (73%), ГТФ (62%), УТФ (16%), ЦТФ (16%) | АТФ (100%), ИТФ (78%), ГТФ (23%), ЦТФ (6%), ТТФ (0,18%) | - | АТФ>ГТФ> ИТФ>УТФ |
Ссылка | Данная работа | [Qiuhao et al., 2004] | [Caescu et al., 2004] | [Fennington, Hughes, 1996] | [Thompson et al., 1991] |
8. Транскрипционная организация генов fruK и ams

 Для изучения потенциальной котранскрипции генов fruK и ams, проводили ПЦР с применением обратной транскриптазы (ОТ-ПЦР). кДНК, синтезированную с праймером As-RT, использовали в качестве матрицы для ПЦР с праймерами lp-as-F и lp-as-R. В результате был получен фрагмент ДНК ожидаемой длины - 1010 п. н. Следовательно, гены fruk и ams сцеплены и транскрибируются в виде одной мРНК (Рис. 11).
Для изучения потенциальной котранскрипции генов fruK и ams, проводили ПЦР с применением обратной транскриптазы (ОТ-ПЦР). кДНК, синтезированную с праймером As-RT, использовали в качестве матрицы для ПЦР с праймерами lp-as-F и lp-as-R. В результате был получен фрагмент ДНК ожидаемой длины - 1010 п. н. Следовательно, гены fruk и ams сцеплены и транскрибируются в виде одной мРНК (Рис. 11).
|
|
|

|
|
|
Одним из продуктов деградации сахарозы под действием амилосахаразы является фруктоза. Котранскрипция генов fruK и ams подтверждает предположение об участии фруктокиназы в реутилизации фруктозы, образовавшейся при катаболизме эндогенной сахарозы.
ЗАКЛЮЧЕНИЕ
В частично аннотированном геноме галотолерантного метанотрофа Methylomicrobium alcaliphilum 20Z нами впервые обнаружен кластер генов sps-spp-fruK-ams, продуктами которых являются сахарозофосфатсинтаза, сахарозофосфатфосфатаза, фруктокиназа и амилосахараза, катализирующие биосинтез и последующий метаболизм сахарозы. Показано, что сахароза накапливается в клетках при увеличении солености среды, но не в ответ на смену источника углерода и энергии с метана на метанол или повышение концентрации метанола в среде. Мутантный штамм, дефектный по генам основного осмопротектора – эктоина, накапливал в 8 раз больше сахарозы, по сравнению с родительским штаммом. Полученные данные однозначно свидетельствуют о том, что Mm. alcaliphilum 20Z синтезирует сахарозу в качестве дополнительного осмопротектора.
Достоверно установлено, что кластер sps-spp-fruK-ams присутствует в геномах как галофильных, так и негалофильных метилотрофов с РМФ путем С1-ассимиляции. Кроме того, гомологи этих генов найдены у галофильных и соленезависимых автотрофных протеобактерий с РБФ-циклом. В совокупности, это указывает на иные, нежели осмопротекторная, функции сахарозы у прокариот. Наличие аналогичных генных кластеров метаболизма сахарозы у различных групп прокариот, а также филогенетический анализ аминокислотных последовательностей соответствующих ферментов свидетельствуют о горизонтальном переносе генов, участвующих в биосинтезе и деградации сахарозы.
Впервые из метанотрофов клонированы гены сахарозофосфатфосфатазы, фруктокиназы и амилосахаразы, получены и охарактеризованы гомогенные препараты этих ферментов. Рекомбинантная сахарозофосфатфосфатаза отличалась от ранее охарактеризованных аналогичных ферментов присутствием нескольких олигомерных форм, а также высокой константой ингибирования сахарозой. Эта особенность сахарозофосфатфосфатазы коррелирует с осмопротекторной ролью сахарозы и необходимостью поддержания относительно высокой внутриклеточной концентрации этого дисахарида.
Рекомбинантная амилосахараза Mm. alcaliphilum 20Z, в отличие от описанных амилосахараз гетеротрофных бактерий, имела на порядок более низкую активность и подчинялась кинетике Михаэлиса-Ментен. Низкая активность амилосахаразы может быть также связана с необходимостью поддерживать определенный пул сахарозы в клетке. Инактивация гена ams приводила к увеличению внутриклеточного содержания сахарозы, что подразумевает участие амилосахаразы в катаболизме дисахарида. Поскольку Mm. alcaliphilum 20Z синтезирует гликогеноподобный полисахарид [Khmelenina et al., 1999], логично предположить, что этот метанотроф реализует путь синтеза гликогена с использованием сахарозы в качестве интермедиата.
Наличие у Mm. alcaliphilum 20Z высокоактивной фруктокиназы, имеющей строгую специфичность и высокое сродство к фруктозе, а также тот факт, что гены fruK и ams котранскрибируются в виде одной матричной РНК, определенно свидетельствуют в пользу участия данного фермента в активировании образующейся фруктозы для последующего включения в метаболизм.
Клонирование и гетерологичная экспрессия гена sps в E. coli не привели к получению активного препарата сахарозофосфатсинтазы. Однако инактивация гена sps сопровождалась потерей способности культуры синтезировать сахарозу, что подразумевает ключевую роль гена этого фермента в биосинтезе сахарозы у Mm. alcaliphilum 20Z. В то же время, наличие гена сахарозофосфатфосфатазы указывает на то, что биосинтез сахарозы у данного метанотрофа происходит двухступенчато, с образованием сахарозо-6-фосфата в качестве интермедиата, т. е. путем, функционирующим у растений и цианобактерий [Page-Sharp et al., 1999; Lunn et al., 2000].
Как известно, Mm. alcaliphilum 20Z реализует РМФ-путь ассимиляции формальдегида с образованием фосфогексоз, в частности, фруктозо-6-фосфата и глюкозо-1-фосфата [Хмеленина с соавт., 1997]. Присутствие в геноме данного метанотрофа генов-гомологов АДФ-глюкопирофосфорилазы (YP004918751.1) и УДФ-глюкопирофосфорилазы (YP004918011.1) свидетельствует о том, что синтез сахарозы происходит именно из этих первичных продуктов РМФ-пути (Рис. 12).
Кроме того, присутствие амилосахаразы и фруктокиназы означает, что эндогенная сахароза в некоторых случаях (например, при понижении солености среды) используется для синтеза осмотически инертного гликогена (Рис. 12). Наличие у Mm. alcaliphilum 20Z гена, кодирующего гликогенсинтазу, свидетельствует о том, что гликоген может синтезироваться альтернативным путем. Из приведенной схемы следует, что при синтезе гликогена с участием сахарозофосфатсинтазы и амилосахаразы затрачивается на одну молекулу нуклеозидтрифосфата больше, чем при участии гликогенсинтазы. Можно предположить, что синтез сахарозы с последующей трансформацией в гликоген служит своеобразным футильным циклом, используемым метанотрофом для сброса избыточной метаболической энергии.
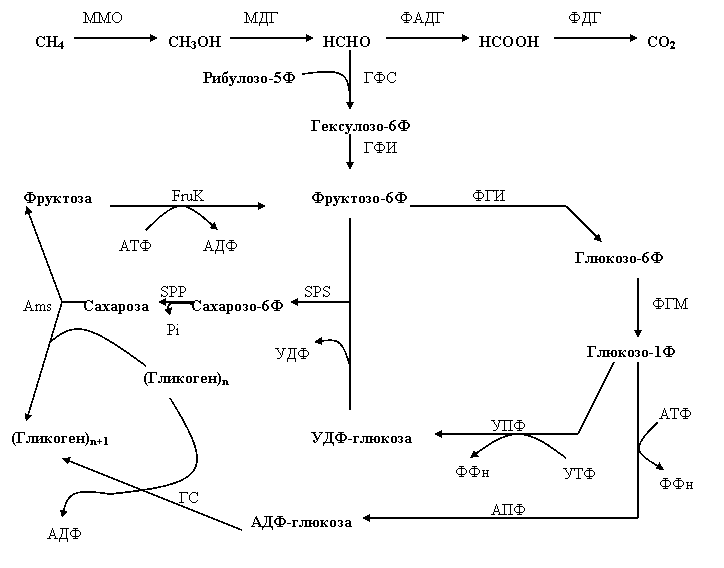

Рис. 12. Предполагаемая схема метаболизма сахарозы у Mm. alcaliphilum 20Z.
MMO – метанмонооксигеназа, МДГ – метанолдегидрогеназа, ФАДГ – формальдегиддегидрогеназа, ФДГ – формиатдегидрогеназа, ГФС – гексулозофосфатсинтаза, ГФМ – гексулозо-фосфатизомераза, ФГИ – фосфоглюкоизомераза, ФГМ – фосфоглюкомутаза, УПФ – УДФ-глюкопирофосфорилаза, АПФ - АДФ-глюкопирофосфорилаза, ГС – гликогенсинтаза, SPS – сахарозофосфатсинтаза, SPP – сахарозофосфатфосфатаза, FruK – фруктокиназа, Ams – амилосахараза.
Таким образом, среди прокариот только фототрофы и хемоавтотрофы, а также аэробные метилотрофы способны синтезировать сахарозу. Исходя из высокой гомологии генов и ферментов метаболизма сахарозы у этих групп бактерий, уместно предположить реализацию у них аналогичного биохимического пути. В совокупности, это подтверждает ранее выдвинутую гипотезу общности происхождения автотрофных и метилотрофных прокариот [Whittenbury et al., 1980].
Данная работа инициировала расшифровку путей метаболизма и функциональной роли сахарозы у метилотрофных протеобактерий. Дальнейшие системные исследования особенностей биологии Mm. alcaliphilum 20Z и негалофильных (Methylobacillus flagellatus KT) или термофильных (Methylocaldum szegediense O-12) метилотрофов, включая анализ их генопротеомов и транскриптомов, приведут к лучшему пониманию феномена полифункциональности сахарозы, а также внесут ясность в сценарий эволюции путей метаболизма сахарозы у прокариот.
ВЫВОДЫ
1 . В геноме галотолерантного метанотрофа Methylomicrobium alcaliphilum 20Z обнаружен кластер из четырех генов, продукты которых – сахарозофосфатсинтаза (SPS), сахарозофосфатфосфатаза (SPP), фруктокиназа (FruK) и амилосахараза (Ams), участвуют в метаболизме сахарозы.
2 . В экспериментах с мутантами доказано, что сахароза является дополнительным к эктоину осмопротектором. Инактивация двух генов пути биосинтеза основного осмопротектора – эктоина, приводит к 8-кратному увеличению накопления сахарозы в клетках мутанта ΔectBC. Впервые полученный инсерционный мутант по гену сахарозофосфатсинтазы (Δsps) не способен накапливать сахарозу, но при высокой солености среды культивирования синтезирует большее количество эктоина, чем исходный штамм.
3 . Установлено, что рекомбинантная сахарофосфатфосфатаза существует в нескольких олигомерных формах с м. м. 33, 66, 132 и 198 кДа, катализирует гидролиз сахарозо-6-фосфата до сахарозы и неорганического фосфата (Vmax = 18,9±0,6 Е/мг, Kmкаж= = 36±4 мкМ).
4 . Обнаружено, что рекомбинантная амилосахараза является мономером с м. м. 75 кДа, катализирует одновременно гидролиз сахарозы (Vmax = 53±1 мЕ/мг, Kmкаж = 11±1 мМ) и трансгликозилирование с переносом глюкопиранозного остатка сахарозы на гликоген (Vmax = 60±2 мЕ/мг, Kmкаж = 6±1 мМ), тем самым участвуя в деградации сахарозы и образовании полисахарида.
5 . Выявлено, что рекомбинантная фруктокиназа – мономер с м. м. 34,5 кДа, катализирует АТФ-зависимое фосфорилирование фруктозы до фруктозо-6-фосфата (Vmax = 141±6 Е/мг, кажущиеся Kmфруктоза = 0,26±0,03 мМ, KmАТФ = 1,3±0,2 мМ). Гены fruK и ams котранскрибируются в виде одной полицистронной мРНК.
6 . Впервые предложена схема путей метаболизма сахарозы у Methylomicrobium alcaliphilum 20Z. Сахароза синтезируется из интермедиатов рибулозомонофосфатного цикла ассимиляции формальдегида: фруктозо-6-фосфата и УДФ-глюкозы. Последующее превращение сахарозы в гликоген, по-видимому, служит не только для запасания углерода, но и для сброса избыточной метаболической энергии в клетках данного метанотрофа.
Список публикаций по теме диссертации:
Статьи в реферируемых журналах, рекомендованных ВАК РФ:
1. , , Троценко рекомбинантной ATP-зависимой фруктокиназы галотолерантного метанотрофа Methylomicrobium alcaliphilum 20Z. Биохимия, 2012, Т. 77, Вып. 4, С. 474 – 480.
2. , , Троценко и характеристика нокаут-мутантов Methylomicrobium alcaliphilum 20Z, дефектных по генам синтеза сахарозы и эктоина. Микробиология. 2013. Т.82. №2. С. 251-253.
Тезисы докладов:
1. , , Решетников генов биосинтеза сахарозы у галотолерантного метанотрофа Methylomicrobium alcaliphilum 20Z. XV Международная Пущинская школа-конференция молодых ученых «Биология - наука XXI века». Пущино, 2011г. С. 360-361.
2. , Решетников и характеристика рекомбинантных фруктокиназы и амилосахаразы из галотолерантного метанотрофа Methylomicrobium alcaliphilum 20Z. Всероссийская конференция с элементами научной школы для молодежи «Экотоксикология-2011». Тула, 2011г. С. 37.
3. , , Решетников сахарозы у галотолерантного метанотрофа Methylomicrobium alcaliphilum 20Z. Всероссийская молодежная школа-конференция «Актуальные аспекты современной микробиологии». Москва, 2011г. С. 7-9.
4. , Решетников и ферменты метаболизма сахарозы у галотолерантного метанотрофа Methylomicrobium alcaliphilum 20Z. XVI Международная Пущинская школа-конференция молодых ученых «Биология - наука XXI века». Пущино, 2012г. С.166-167.
5. , Решетников и ферменты метаболизма сахарозы у галотолерантного метанотрофа Methylomicrobium alcaliphilum 20Z. Всероссийская молодежная школа-конференция «Актуальные аспекты современной микробиологии». Москва, 2012г. С. 7-9.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
