
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Идентичные кластеры генов, кодирующих предполагаемые ферменты метаболизма сахарозы (sps-spp-pfkB-ams), обнаружены нами у других представителей метанотрофов (Methylomicrobium album BG8 и Methylobacter tundripaludum SV96), а также у не растущих на метане метилобактерий (Methylophaga aminisulfidivorans MP и Methylophaga thiooxydans DMS010). У Methylomonas methanica MC09, в отличие от других метанотрофов, ген амилосахаразы расположен дистантно, т. е. отдельно на хромосоме. Кроме того, кластеры sps-spp-pfkB-ams обнаружены у цианобактерии Synechococcus sp. PCC 7002 и хемоавтотрофных протеобактерий (Mariprofundus ferrooxydans PV-1, Thiocapsa marina 5811, Thiomicrospira crunogena XL-1 и Marichromatium purpuratum 984 (Рис. 4).
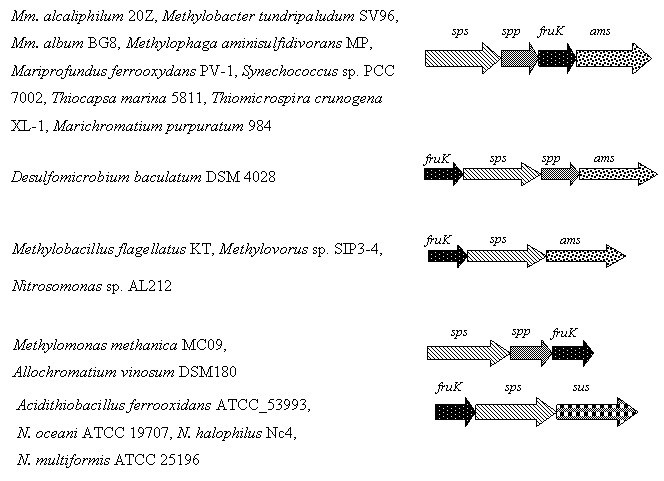 |
Рис. 4. Структура кластеров генов, кодирующих предполагаемые ферменты метаболизма сахарозы у метилотрофов, хемоавтотрофов и цианобактерий.
У некоторых бактерий структура генного кластера метаболизма сахарозы отличалась от таковой Mm. alcaliphilum 20Z. Так, у галофильных аммоний-окисляющих Nitrosococcus halophilus Nc4 и Nitrosococcus oceani ATCC 19707 гены spp располагаются отдельно на хромосоме, хотя сахарозофосфатфосфатазы этих бактерий филогенетически близки ферменту из Mm. alcaliphilum 20Z (41,3% и 40,1% идентичности, соответственно).
Более того, в хромосоме метилобактерий Mb. flagellatus KT и Methylovorus sp. SIP3-4, а также у хемовтотрофов A. ferrooxidans ATCC_53993 и Nitrosospira multiformis ATCC 25196 отсутствует ген spp. Интересно, что С-концевой домен как растительных, так и большинства цианобактериальных сахарозофосфатсинтаз гомологичен сахарозофосфатфосфатазе [Cumino et al., 2002; Lunn, 2002; Lunn and MacRae, 2003]. На этом основании предположено, что фермент может катализировать как синтазную, так и фосфатазную реакции [Lunn, 2002].
Итак, анализ доступных в Database геномов микроорганизмов выявил у хемоавтотрофных и метилотрофных бактерий присутствие генов, продукты которых проявляют гомологию с ферментами биосинтеза и реутилизации сахарозы, и которые, как правило, локализованы в трех - или четырехгенном кластерах. Однако гены и ферменты метаболизма сахарозы у протеобактерий не изучались. К настоящему времени охарактеризованы только сахарозофосфатсинтазы и сахарозофосфатфосфатазы некоторых растений и цианобактерий, а ферменты утилизации экзогенной сахарозы – только у бактерий, использующих дисахарид в качестве ростового субстрата. В этой связи представлялось интересным определить функциональность генов кластера sps-spp-pfkB-ams у Mm. alcaliphilum 20Z, выделить и охарактеризовать соответствующие ферменты.
3. Определение функциональной роли гена сахарофосфатсинтазы (sps)
Ген sps (GenBank CCE22309.1) амплифицировали с геномной ДНК Mm. alcaliphilum 20Z и клонировали в векторе pET30a+, далее рекомбинантной плазмидой pETsps трансформировали компетентные клетки E. coli Rosetta (DE3). Экспрессию рекомбинантного белка SPS-His6 индуцировали добавлением ИПТГ. Фермент был очищен из лизата клеток E. coli до электрофоретически гомогенного состояния методом аффинной металл-хелатной хроматографии на колонке с Ni2+-NTA-агарозой. Однако полученный препарат фермента был неактивен. Для определения функциональной роли sps нами был получен мутант с инсерцией канамициновой кассеты в данном гене по схеме мутагенеза, аналогичной таковой для конструирования мутанта ectBC- (Рис. 2). В клетках мутанта, выращенных на метаноле (0,5% по объему) при различной солености среды, определяли содержание сахарозы. Обнаружено, что полученный мутант не накапливал сахарозу. Это свидетельствовало о ключевой роли продукта гена sps в биосинтезе сахарозы у Mm. alcaliphilum 20Z, а также о том, что у данного метанотрофа функционирует единственный путь синтеза сахарозы с участием сахарозофосфатсинтазы.
4. Характеристика рекомбинантой сахарозофосфатфосфатазы (SPP).
![]()
![]()
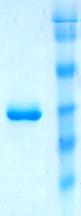
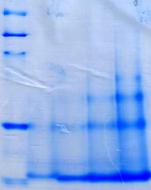
![]() Ген spp (GenBank CCE22310.1) амплифицировали с геномной ДНК Mm. alcaliphilum 20Z и клонировли в вектор pET28b. Фермент, экспрессированный в клетках E. coli Rossetta (DE3), очищали аффинной металл-хелатной хроматографией до электрофоретически гомогенного состояния. Согласно данным ДСН-ПААГЭ, электрофоретическая подвижность белка соответствовала м. м. ~35 кДа (Рис. 5А), что согласуется с теоретически рассчитанной м. м. (32,7 кДа). Нативный электрофорез в ПААГ с градиентом пористости выявил несколько белковых полос с м. м. примерно 35, 70, 150 и 210 кДа, что указывает на возможность существования фермента одновременно в форме мономера, димера, тетрамера и гексамера (Рис. 5Б)
Ген spp (GenBank CCE22310.1) амплифицировали с геномной ДНК Mm. alcaliphilum 20Z и клонировли в вектор pET28b. Фермент, экспрессированный в клетках E. coli Rossetta (DE3), очищали аффинной металл-хелатной хроматографией до электрофоретически гомогенного состояния. Согласно данным ДСН-ПААГЭ, электрофоретическая подвижность белка соответствовала м. м. ~35 кДа (Рис. 5А), что согласуется с теоретически рассчитанной м. м. (32,7 кДа). Нативный электрофорез в ПААГ с градиентом пористости выявил несколько белковых полос с м. м. примерно 35, 70, 150 и 210 кДа, что указывает на возможность существования фермента одновременно в форме мономера, димера, тетрамера и гексамера (Рис. 5Б)![]()
![]()
![]()
![]()
![]()
![]()
|
Рекомбинантный фермент катализировал гидролиз сахарозо-6-фосфата до сахарозы и неорганического фосфата. Наибольшую активность сахарозофосфатфосфатаза проявляла при 35ºС и рН 6,5 (Табл.1).
Интересно отметить, что оптимумы работы SPP не совпадали с оптимальными условиями роста штамма 20Z (рН 9 и 30oC), но близки таковым у сахарозофосфатфосфатаз из других организмов (Табл. 1). Возможно, это объясняется приобретением генов синтеза сахарозы путем горизонтального переноса от мезофилов.
Зависимость активности SPP от концентрации сахарозо-6-фосфата соответствовала классическому уравнению Михаэлиса-Ментен (Vmax = 18,9±0,6; Km = 36±4 мкМ). Активность сахарофосфатфосфатазы незначительно ингибировалась сахарозой (Ki = 1±0,6 М). Данная константа ингибирования гораздо выше, чем у других охарактеризованных SPP. По-видимому, эта особенность фермента из Mm. alcaliphilum 20Z обусловлена необходимостью поддержания высокой внутриклеточной концентрации сахарозы.
Таблица. 1. Свойства некоторых сахарозофосфатфосфатаз
Параметры | Mm. alcaliphilum 20Z | Synechocystis sp.PCC 6803 | Anabaena sp. PCC 7120 | Oryza sativa | Pisum sativum |
Молекулярная масса субъединицы, кДа | 32,7 | 27 | 28 | 50 | 55 |
Субъединичная структура | мономер, димер, тетрамер, гексамер | мономер | мономер | димер | димер |
pH оптимум | 6,5 | 6,8 | 6,5 | 6,5 | 6,8 |
Температурный оптимум (oC) | 35 | - | - | - | - |
Km (мМ) | 0,036 | 0,0075 | 0,35 | 0,065 | 0,25 |
Vmax (Е/мг) | 18,9 | 46 | 15,7 | 1250 | - |
Ki (сахароза) (мМ) | 1000 | 161 | 80 | - | - |
Ссылка | Данная работа | [Lunn, 2002] | [Cumino et al., 2001] | [Lunn et al., 2000] | [Whitaker, 1984] |
5. Характеристика рекомбинантой амилосахаразы (Ams).
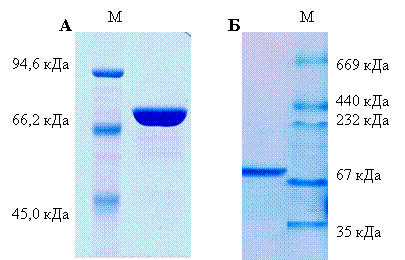 Ген ams (GenBank CCE22312.1) амплифицировали с геномной ДНК Mm. alcaliphilum 20Z и клонировали в вектор pET30a+. Рекомбинантный фермент Ams-His6 экспрессировали в клетках E. coli Rossetta (DE3), очищали аффинной металл-хелатной хроматографией до электрофоретически гомогенного состояния. Электрофоретическая подвижность белка в ДСН-ПААГ соответствовала ~75 кДа (Рис. 6А), что согласуется с расчетной м. м. (76,7 кДа). Нативный электрофорез в градиенте ПААГ (4-30%) выявил единственную полосу с м. м. около 70 кДа (Рис. 6Б), что свидетельствует о мономерной форме фермента.
Ген ams (GenBank CCE22312.1) амплифицировали с геномной ДНК Mm. alcaliphilum 20Z и клонировали в вектор pET30a+. Рекомбинантный фермент Ams-His6 экспрессировали в клетках E. coli Rossetta (DE3), очищали аффинной металл-хелатной хроматографией до электрофоретически гомогенного состояния. Электрофоретическая подвижность белка в ДСН-ПААГ соответствовала ~75 кДа (Рис. 6А), что согласуется с расчетной м. м. (76,7 кДа). Нативный электрофорез в градиенте ПААГ (4-30%) выявил единственную полосу с м. м. около 70 кДа (Рис. 6Б), что свидетельствует о мономерной форме фермента.
Рис. 6. 8% ДСН-ПААГ электрофорез (А) и нативный градиентный 4-30% ПААГЭ рекомбинантной амилосахаразы.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
