
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Аналогично другим описанным амилосахаразам, рекомбинантная Ams из Mm. alcaliphilum 20Z катализировала одновременно гидролиз сахарозы до фруктозы и глюкозы, а также трансгликозилирование с переносом глюкопиранозного остатка сахарозы на полигликан [van der Veen et al., 2006; Pizzut-Serin et al., 2005; Jung et al., 2009; Ha et al., 2009]. Фермент проявлял максимальную активность при 30oC и рН 8,0, что согласуется с экофизиологическими параметрами, оптимальными для роста Mm. alcaliphilum 20Z.
Зависимость активности рекомбинантной амилосахаразы Mm. alcaliphilum 20Z от концентрации сахарозы подчинялась классической кинетике Михаэлиса-Ментен (Рис. 7), в отличие от ранее описанных Ams из Neisseria polysaccharea [De Montalk et al., 2000] и Deinococcus radiodurans [Pizzut-Serin et al., 2005]. Амилосахаразы этих гетеротрофных бактерий имели два плато на кривой зависимости активности от концентрации сахарозы и, соответственно, по два значения Vmax и Km (Табл. 3). У амилосахаразы из N. polysaccharea и D. radiodurans имеется дополнительный сайт связывания сахарозы (SB2), наличие которого, по-видимому, обуславливает необычные кинетические свойства фермента [Skov et al., 2002]. Выравнивание аминокислотных последовательностей амилосахараз показало, что у Ams из Mm. alcaliphilum 20Z в позиции, соответствующей Gln445 и Tyr446 в SB2 сайте N. polysaccharea, находятся метионин и аргинин (Рис. 7), что, вероятно, делает дополнительный субстрат-связывающий сайт нефункциональным.
Mm. alcaliphilum : DSIASGVPFMRNEKTGDARIAGSLA :
Mm.album : HSPARGLVFMRNDETGDARICGSLA :
Methylovorus : DSIARGVAFGRNDKTGDARISGSLA :
Mb.flagellatus : GSTATGVAFGVNPKTGDARISGSLA :
Mp.aminisulfidivorans : HSHARGLPFGENEKTGDARISGSLA :
Synechococcus sp.7002 : DT-SRGLEFMRNDSTGDARICGSLA :
T. crunogena : ESHARGLPFGENAKTGDARISGSLA :
N.polysaccharea :GSFARGVPFQYNPSTGDCRVSGTAA :
N.subflava : GSFARGVPFQYNPSTGDCRVSGTAA :
D.geothermalis : GSFARGLVFQYNPVNGDRRISGSAA :
D.radiodurans : GSFARGLVFQHHPQTGDRRISGTAA :
![]() Рис. 7. Выравнивание аминокислотных последовательностей амилосахараз. Рамкой обозначены аминокислоты, участвующие в связывании сахарозы в дополнительном субстрат-связывающем сайте амилосахаразы из N. polysaccharea [Skov et al., 2002].
Рис. 7. Выравнивание аминокислотных последовательностей амилосахараз. Рамкой обозначены аминокислоты, участвующие в связывании сахарозы в дополнительном субстрат-связывающем сайте амилосахаразы из N. polysaccharea [Skov et al., 2002].
Амилосахараза имела более высокое сродство к сахарозе в реакции трансгликозилирования (табл. 2), однако при введении в реакционную смесь затравки гликогена кажущаяся Km для сахарозы как в реакции гидролиза, так и в реакции трансгликозилирования была одинакова. Кроме того, добавление гликогена увеличивало скорость трансгликозилирования, при этом скорость гидролиза сахарозы не изменялась.
Исходя из полученных результатов, можно предположить, что основной функцией амилосахаразы является перевод избыточного количества эндогенной сахарозы Mm. alcaliphilum 20Z в полигликаны типа гликогена.
Таблица 2. Кинетические параметры рекомбинантной амилосахаразы Mm. alcaliphilum 20Z.
Реакция | Без добавления гликогена | В присутствии 0,1 мг/мл гликогена | ||
Vmax, нмоль/(мин*мг) | Kmкаж, мМ | Vmax, нмоль/(мин*мг) | Kmкаж, мМ | |
Трансгликозилирование | 60±2 | 6±1 | 98±1 | 11,2±0,8 |
Гидролиз | 53±1 | 11±1 | 51±1 | 12±1 |
6. Получение инсерционного мутанта по гену ams
Методом двойной гомологичной рекомбинации был получен мутант с инсерцией канамициновой кассеты в гене ams.
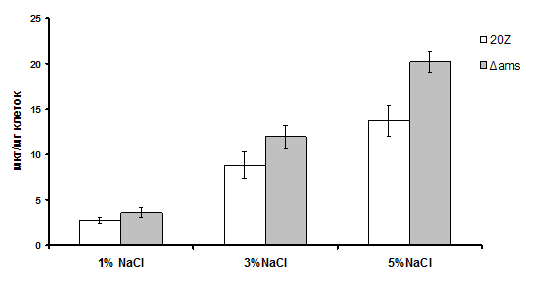
Рис. 8. Накопление сахарозы клетками штаммов 20Z и Δams, выращенных при различном содержании NaCl в среде.
Полученный мутант и штамм дикого типа выращивали на метаноле (0,5% по объему) при различных концентрациях NaCl. В клетках мутанта обнаружено примерно в 1,3 раза больше сахарозы, по сравнению с исходным штаммом (Рис. 8), что подтверждает участие амилосахаразы в деградации эндогенной сахарозы у Mm. alcaliphilum 20Z.
Таблица 3. Свойства некоторых амилосахараз прокариот.
Параметр | Mm. alcaliphilum | Neisseria polysaccharea | Deinococcus radiodurans | Deinococcus geothermalis | Alteromonas addita |
Субъединичная структура | мономер (1×76 кДа) | мономер (1×70 кДа) | |||
pH оптимум | 8,0 | 6,0 | 7,0 | 7,0 (Гидролиз) 8,0 (Трансгликозилирование) | |
Температурный оптимум, oC | 30 | 42 | 36 | 47 | 45 |
Km, мМ | |||||
Общая активность | 8,1 (11,3)* | 4,0 ([сахароза] < 20 мМ) 71 ([сахароза] > 20 мМ) | 10 ([сахароза] < 41 мМ) 84 ([сахароза] > 41 мМ) | ||
Трансгликозилирование | 6 (11,2)* | 8,1 ([сахароза] < 20 мМ) 112 ([сахароза] > 20 мМ | |||
Гидролиз | 11 (11)* | 2,5 ([сахароза] < 20 мМ) 29 ([сахароза] > 20 мМ) | |||
kcat, мин-1 | |||||
Общая активность | 8,7 (11,2)* | 44([сахароза] < 20 мМ) 84 ([сахароза] > 20 мМ) | 49,2 ([сахароза] < 41 мМ) 121 ([сахароза] > 41 мМ) | ||
Трансгликозилирование | 4,6 (7,5)* | 26 ([сахароза] < 20 мМ) 54 ([сахароза] > 20 мМ) | |||
Гидролиз | 4,1 (4,0)* | 21 ([сахароза] < 20 мМ) 31 ([сахароза] < 20 мМ) | |||
Ссылка | Данная работа | [Jung et al., 2009; van der Veen et al., 2006] | [Emond et al., 2008; Pizzut-Serin et al., 2005] | [Jung et al., 2009] | [Ha et al., 2009] |
* - в присутствии 0,1 мг/мл гликогена
7. Характеристика рекомбинантой фруктокиназы (FruK).
Ген, предварительно аннотированный как pfkB (GenBank CCE22311.1), амплифицировали с геномной ДНК Mm. alcaliphilum 20Z и клонировали в вектор pET22b+. Рекомбинантный фермент экспрессировали в клетках E. coli Rossetta (DE3), трансформированных плазмидой pETfruK. Фермент очищали из лизата клеток аффинной металл-хелатной хроматографией на колонке с Ni2+-NTA-агарозой до электрофоретически гомогенного состояния. Молекулярная масса субъединицы, определенная с помощью ДСН-ПААГЭ, соответствовала расчетной (34,5 кДа) (Рис. 9А). Нативный градиентный (4-30%) электрофорез в ПААГ выявил единственную полосу, соответствующую м. м. около 35 кДа (Рис. 9Б). Резонно предположить, что фермент in vivo является мономером.
![]()
![]() Рекомбинантный белок катализировал АТФ-зависимое фосфорилирование фруктозы до фруктозо-6-фосфата, т. е. являлся фруктокиназой. Рекомбинантная FruK имела абсолютную специфичность к фруктозе, как акцептору фосфорильной группы, но не проявляла активности с глюкозой, ксилозой, рибозой, арабинозой, мальтозой, сахарозой, а также с глюкозо-6-фосфатом и фруктозо-6-фосфатом. Фруктокиназа была способна использовать в качестве фосфорильного донора АТФ, ГТФ и УТФ, но не ЦТФ и не пирофосфат.
Рекомбинантный белок катализировал АТФ-зависимое фосфорилирование фруктозы до фруктозо-6-фосфата, т. е. являлся фруктокиназой. Рекомбинантная FruK имела абсолютную специфичность к фруктозе, как акцептору фосфорильной группы, но не проявляла активности с глюкозой, ксилозой, рибозой, арабинозой, мальтозой, сахарозой, а также с глюкозо-6-фосфатом и фруктозо-6-фосфатом. Фруктокиназа была способна использовать в качестве фосфорильного донора АТФ, ГТФ и УТФ, но не ЦТФ и не пирофосфат.
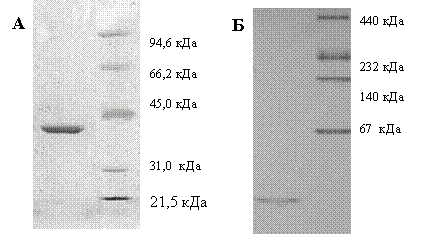 |
Рис. 9. 12% ДСН-ПААГ электрофорез (А) и градиентный 4-30% ПААГ электрофорез (Б) рекомбинантной фруктокиназы.
Активность фермента зависела от ионов двухвалентных металлов (относительная активность): Mg2+ (100%), Mn2+ (146%), Co2+ (63%), Zn2+(1,1%), Ni2+ (0,8%). Активность фермента не обнаруживалась в отсутствии металла-кофактора. Максимальная активность проявлялась при соотношении концентраций АТФ и Mg2+ в диапазоне 1:1 – 1:1,5 (Рис. 10). По-видимому, фруктокиназа использует комплекс Mg-АТФ и ингибируется свободным АТФ и высокими концентрациями ионов Mg2+.
Рекомбинантная фруктокиназа была максимально активна при рН 9,0, что коррелирует с оптимальным значением рН для роста штамма 20Z. Напротив, температурный оптимум FruK (60ºС) вдвое превышал таковой для роста данного метанотрофа. Аномально высокий температурный оптимум был обнаружен также у фруктокиназы из Bifidobacterium longum [Caescu et al., 2004] (Табл. 4).
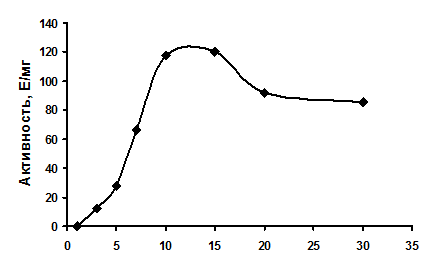
Рис. 10. Зависимость активности фруктокиназы от концентрации ионов Mg2+ при постоянной концентрации АТФ (10 мМ).
Фруктокиназа из Mm. alcaliphilum 20Z подчинялась классической кинетике Михаэлиса-Ментен, имела достаточно высокую активность и высокое сродство к фруктозе (Vmax = 141±6 Е/мг, кажущиеся Kmфруктоза = 0,26±0,03 мМ, KmАТФ = 1,3±0,2 мМ), что указывает на способность данного метанотрофа метаболизировать фруктозу при достаточно низких концентрациях в клетке. Поскольку Mm. alcaliphilum 20Z не растет на фруктозе или других углеводных субстратах, роль фруктокиназы у данного метанотрофа не связана с ассимиляцией экзогенных сахаров. Учитывая расположение гена fruK в кластере генов метаболизма сахарозы, можно предположить участие фруктокиназы в реутилизации сахарозы, образующейся при катаболизме эндогенной сахарозы амилосахаразой.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
