

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Следует отметить, что многие экспериментальные данные получены именно в предположении простейшей модели связывания, когда молекулы лиганда и рецептора рассматриваются как свободные и моновалентные. В действительности взаимодействие является мультивалентным. К тому же, в реальной биологической системе взаимодействие вирус-клетка, как правило, происходит в условиях, когда один из реагентов является иммобилизованным на поверхность либо культуральной посуды, либо в тканях организма, что является случаем гетерогенного связывания.
При моделировании мультивалентного связывания наибольшее распространение получило «приближение эквивалентных мест связывания» («equivalent site approximation»), в котором места для связывания лиганда полагаются идентичными и взаимодействующими с различными рецепторами независимо [36]. При этом пренебрегается возможными внутримолекулярными реакциями, которые, например, могут иметь место для мультивалентных рецепторов [37].
При таком подходе мультивалентный лиганда с валентностью n может связаться с поверхностью клетки i=1,…,n раз. Анализ в случае наличия кооперативного связывания (в случае сильно выраженной положительной кооперативности) может быть сильно упрощен [38], если предположить, что кооперативное лиганд-рецепторное взаимодействие описывается схемой
![]()
Даже в равновесии и в случае очень сильных упрощающих предположений, уравнения на концентрации реагентов достаточны сложны.
Значения константы скорости ассоциации комплекса антиген-антитело опубликованные в научной литературе, колеблются от 106 до 108 М-1с-1. В то время как значения для константы скорости диссоциации комплекса антиген-антитело варьируются в больших пределах от 10-4 до 10000 с-1 [39]. Такая разница в разбросе значений для констант скоростей прямой и обратной реакций ведет к предположению, которое на сегодняшний день является общепризнанным взглядом: константы скорости прямой реакции связывания отличаются для разных молекул антигена и антитела несильно, а имеющиеся различия в константах связывания обусловлены в основном различиями в константе скорости диссоциации. Выражение для равновесной константы диссоциации легко получить из уравнения ![]() (учитывая (1.1)):
(учитывая (1.1)):
Особенностью методического подхода, реализованного в данной работе, является определение кинетических параметров вирус - клеточной системы с помощью детекции изменения pH при образовании вирус-рецепторного комплекса (необратимой адсорбции) и при проникновении вирусной частицы в эндосому клетки.
- Цель работы
Целью данной работы является изучение кинетики активации клетки в результате рецептор-обусловленного эндоцитоза вирусной частицы. В ходе выполнения работы решались следующие задачи:
o Разработка нового метода исследования вирус-клеточного взаимодействия с применением флуоресцентной детекции
o Построение теоретической модели и разработка методики нахождения кинетических параметров вирус-клеточного взаимодействия и определения эффективности ингибитора с применением флуоресцентной детекции
o Применение построенной теоретической модели для аппроксимации результатов кинетического эксперимента
o Сравнение стандартных вирусологических (ФОЕ и ИФА) и нового флуоресцентного методов оценки эффективности ингибитора
Теоретическая часть
- Кинетическая модель связывания вируса с клеткой
(соответствие с условиями проведения эксперимента при 4˚С)
Отличительной особенностью взаимодействия вируса с клеткой является фазовая гетерогенность системы. При связывании вируса гриппа с культурой клеток (моновалентная модель) схема реакции имеет вид, схожий со схемой связывания вируса гриппа с аналогом клеточного рецептора (белок фетуин):
![]()
где [V]- концентрация свободного вируса в суспензии
[R] – концентрация свободных рецепторов на поверхности клеток
[VR] –концентрация вируса на поверхности клетки, связанного с клеточными рецепторами
[V0,] [R0] – начальная концентрация вирусных частиц и клеточных рецепторов соответственно
k+, k - - кинетические константы связывания вируса гриппа с клеткой: ассоциации и диссоциации соответственно
kin - константа проникновения вируса внутрь клетки (константа эндоцитоза)
![]()
где 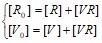 (закон сохранения вещества для вирусных частиц и для рецепторов на поверхности клетки)
(закон сохранения вещества для вирусных частиц и для рецепторов на поверхности клетки)
Приближения:
- [R]>>[VR] ;
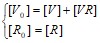 ;
; тогда дифференциальное уравнение примет вид:
![]()
откуда можно найти скорость изменения концентрации вируса на поверхности клетки:
![]()
Таким образом, концентрация вируса на поверхности клетки зависит как и от начальной концентрации вируса так и от концентрации рецепторов на поверхности клетки. А скорость насыщения зависит от начальной концентрации вируса в суспензии. На временах ![]() наступает насыщение клеточных рецепторов вирусом (уже со сделанной оговоркой, что [V0]<<[R0]) Т. е. на таких временах связывание вируса с клеточными рецепторами можно рассматривать как равновесный процесс.
наступает насыщение клеточных рецепторов вирусом (уже со сделанной оговоркой, что [V0]<<[R0]) Т. е. на таких временах связывание вируса с клеточными рецепторами можно рассматривать как равновесный процесс.
- Кинетическая модель процесса эндоцитоза
(соответствие с условиями проведения эксперимента при 37˚С)
Одним из этапов заражения вирусом клетки является процесс эндоцитоза. Рассмотрим его в рамках кинетической модели. Этому посвящен ряд статей [39-40] и их модели заслуживают отдельного рассмотрения, но все они отличаются сложностью в обработке и расчете параметров взаимодействия вируса с живой клеткой (и имеют ряд приближений на этом этапе - этапе расчета параметров взаимодействия вируса с живой клеткой). Мы же попробуем создать упрощенную модель связывания вируса гриппа с живой клеткой, которая не будет требовать приближенных методов анализа и будет адекватно описывать связывание вируса с рецепторами и проникновение вируса внутрь. Поскольку только вирус, связанный с поверхностью клетки, способен проникнуть внутрь клетки, то разумно весь процесс первого этапа заражения вирусом клетки разбить на две последовательные реакции: связывание вируса с клеточными рецепторами и проникновение связанного вируса внутрь клетки (процесс эндоцитоза), или в виде схемы:
где [V]- концентрация свободного вируса в суспензии
[R] – концентрация свободных рецепторов на поверхности клеток
[VR]s/in –концентрация вируса на поверхности/внутри клетки, связанного с клеточными рецепторами
[V0,] [R0] – начальная концентрация вирусных частиц и клеточных рецепторов соответственно
k+, k - - кинетические константы связывания вируса гриппа с клеткой: ассоциации и диссоциации соответственно
kin - константа проникновения вируса внутрь клетки (константа эндоцитоза)
Получаем систему уравнений, моделирующую связывание вируса с клеткой и проникновение вируса в клетку:
Приближения:
- [R]>>[VR]S, [R]>>[V];
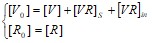 , подставляем в систему (2.6), получаем:
, подставляем в систему (2.6), получаем: Принято (как и ранее), что концентрация (соответственно и количество) свободных рецепторов не изменяется, что является следствием экспериментального ограничения: концентрация вируса много меньше концентрации рецепторов.
Если сделать соответствующую замену:
![]()
то систему дифференциальных уравнений первого порядка можно свести к дифференциальному уравнению второго порядка:
![]()
где
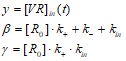
Это уравнение решается и записывается в виде суммы общего решения yo и частного yч: y= yo+ yч
Составим характеристическое уравнение (т. е. ищем решение в виде ![]() ):
):
![]()
Решения этого квадратного уравнения имеют вид:
![]()
Поэтому
![]()
И решение неоднородного уравнения (2.7):
![]()
или для концентрации вирусных частиц на поверхности клетки [VR]s(t):
![]()
Поскольку в начальный момент времени концентрация вирусных частиц на поверхности клетки, а следовательно, и внутри клетки, равна нулю, то, подставляя эти значения в [VR]s(t), получим:
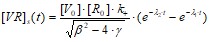
Данная зависимость представляет собой зависимость концентрации вируса на поверхности клетки от концентраций связывающихся веществ и от кинетических характеристик связывания вируса с клеткой.

От начальной концентрации вируса зависит только величина максимального пика, а скорости (сама кинетика) зависят от кинетических параметров и начальной концентрации свободных рецепторов. Видно, что при отсутствии эндоцитоза (γ=0) формула переходит в уже известную (2.4).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
