

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

На данном рисунке 11 детектируемый сигнал - сумма интенсивностей на поверхности и внутри клетки (с соответствующим коэффициентом b=0,4) и есть решение уравнения (3.12).
Экспериментальная часть
Одной из основных задач данной дипломной работы является изучение взаимодействия вируса гриппа именно с живой клеткой. Живая система характеризуется своей изменчивостью во времени, она постоянно находиться в движении. Поэтому при проведении экспериментов очень важно следить за более или менее одинаковыми внешними и внутренними условиями эксперимента. Например, очень важно следить за концентрацией вирусных частиц в суспензии: это влияет не только на определение параметров связывания вируса с клеткой, но и на применимость самой модели (должно соблюдаться условие [R]>>[V], см. Теоретическую часть). Главной изменяющейся единицей в этом процессе является клетка, а точнее ее рецепторы.
При проведении экспериментального изучения кинетики взаимодействия вируса с живой культурой клеток предварительно были охарактеризованы следующие параметры:
оптимальный возраст монослоя клеток, при котором рецепторные свойства клеток меняются незначительно за время проведения кинетического эксперимента (~48 часов [41]) оптимальная концентрация клеток и, как следствие, концентрации клеточных рецепторов(R=N∙r, N – количество клеток в монослое, r – количество рецепторов на поверхности одной клетки) (~106 клеток в лунке) концентрация вирусных частиц в суспензии, используемой для эксперимента, (~10-13M, ~27 по ГА (гемаглютинирующая активность))- Данные, полученные методом ФОЕ
Как уже было упомянуто в разделе Материалы и Методы, в основе методики ингибирования фокусообразующих единиц лежит подвление количества фокусов (зараженных вирусом гриппа штамма A/Aichi/2/68 монослоя клеток MDCK) различными концентрациями аналога клеточных рецепторов – блокатора фетуина. Методика пробит – анализа позволила получить необходимый параметр эффективности данного ингибитора на основе полученных экспериментальных зависимостей «Доза ингибитора - ингибирующий эффект».
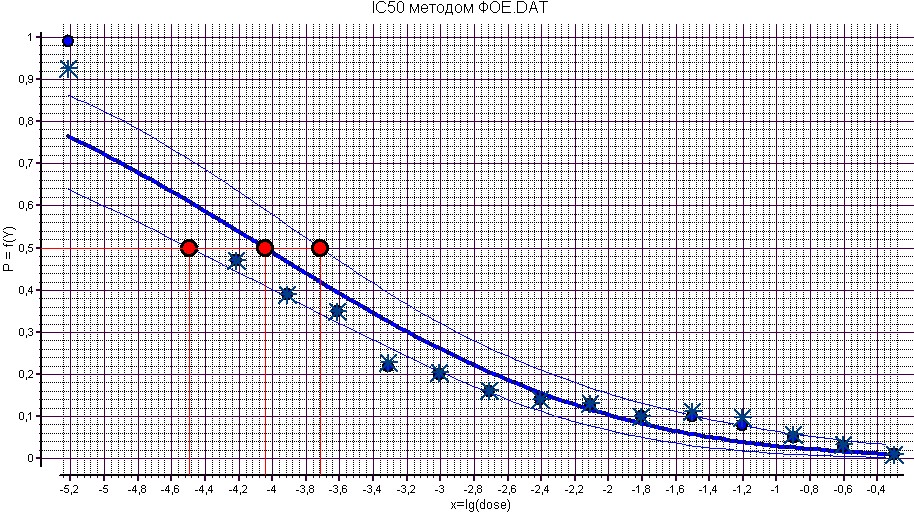

На рисунке 12 представлена зависимость процента подавления заражения культуры клеток MDCK вирусом гриппа штамма A/Aichi/2/68 от логарифма дозы ингибитора фетуин. Рассчитанный IC50 составляет ![]() М.
М.
- Данные метода иммуноферментного анализа (ИФА) [41]
На рис.13 представлено взаимодействие штаммов вируса гриппа с аналогом клеточного рецептора белком фетуином (при +100 С) как в виде экспериментальных точек, так и теоретическое, представляющее собой прямую линию, проведенную через экспериментальные точки в координатах Скэтчарда, согласно уравнению:
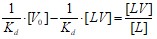

Тангенс угла наклона (рисунок 13) характеризует равновесную константу ассоциации. Т. е. чем больше угол наклона прямой, тем меньше константа ассоциации, соответственно больше константа диссоциации. Из данных, приведенных на рисунке 13, получим константу взаимодействия вируса гриппа штамма A/Aichi/2/68 с аналогом клеточных рецепторов белком фетуин (при +100С):
Kd= (0,64±0,1)·10-10М.
Далее аналогичным способом рассматриваются кинетики взаимодействия вируса с культурой клеток линии MDCK при +320С и +100С по данным которых мы сможем определить кинетические параметры, необходимые для решения обратной задачи (рисунки 14,15)


Исследуя зависимости, приведенные на рисунках 14 и 15 были получены кинетические параметры, приведенные в таблице 1
Таблица 1 Кинетические параметры, полученные методом ИФА
Т, 0С | Kd, M | k+, M-1·s-1 | k-, s-1 | kin, s-1 |
+10 | (3,1±0,3)·10-11 | (3,3±0,3)·104 | (1±0,1)·10-4 | 0 |
+32 | (8±0,6)·10-11 | (1±0,1)·105 | (3±0,4)·10-4 | (2±0,2)·10-4 |
Данные параметры позволят более адекватно подобрать аппроксимацию для решения обратной задачи флуоресцентной детекции, а также позволят оценить степень достоверности результатов полученных методом флуоресцентной детекции.
- Данные, полученные методом флуоресцентной детекции
Для более быстрой оценки IC50 и для более тщательного исследования вирус-клеточного взаимодействия мы предлагаем новый метод – флуоресцентной детекции. Суть его заключается в детекции изменений, происходящих на поверхности и внутри клетки во время процесса эндоцитоза (подробнее – Материалы и Методы).
Для этого мы использовали лизо-сенсор DND–167, интенсивность флуоресценции которого резко увеличивается при понижении pH в диапазоне от 6, до 5. Этот же диапазон соответствует изменению pH на поверхности и в эндосоме клетки при проникновении вируса. Таким образом, наблюдая изменение интенсивности флуоресценции окрашенного вируса на поверхности и внутри клетки, мы можем судить о стадии процесса эндоцитоза в текущий момент времени.

На рисунке 16 изображена зависимость интенсивности флуоресценции от времени, прямопропорциональная концентрации вируса, участвующего в процессе эндоцитоза (3.6). На первых 10 минутах взаимодействия вируса с клеточными рецепторами виден пик флуоресценции, что говорит о начале адсорбции вируса на поверхность клеток (в данном случае DND-167 активируется за счет понижения pH в местах связывания вируса с клеткой, происходящего в результате конформационных изменений HA). Далее, интенсивность не спадает до нуля, а выходит на некоторый уровень, что свидетельствует о постепенном проникновении вируса в клетку (здесь начинается активация лизо-сенсора DND-167 за счет снижения pH в эндосоме клетки). Данную интерпритацию подтверждает та же зависимость, но для ингибированного фетуином (концентрация ~3·10-8М) вируса: пик флуоресценции сдвигается в район 16 минут, что говорит о торможении процесса адсорбции за счет введения конкурента; общая интенсивность падает ~ в 5 раз, что говорит о торможении всего процесса эндоцитоза и постепенной остановке проникновения вируса в клетку.
1) Нахождение параметра эффективности ингибитора IC50
Исследуя зависимость интенсивности сигнала флуоресценции после 20 минут взаимодействия вируса с клеточной суспензией от концентрации блокатора фетуина, стало очевидно, что чем больше концентрация ингибитора, тем сильнее подавляется сигнал флуоресценции. Таким образом был проведен расчет IC50 методом пробит-анализа на основе полученных экспериментальных зависимостей «Доза ингибитора - ингибирующий эффект» используя результаты зависимости подавления сигнала флуоресценции от концентрации блокатора фетуина.

На рисунке 17 представлена зависимость доли подавления заражения культуры клеток MDCK вирусом гриппа штамма A/Aichi/2/68 от логарифма дозы ингибитора фетуин.
Рассчитанный IC50 составляет ![]() М.
М.
2) Применение построенной теоретической модели на кинетическом эксперименте
Расчет теоретической зависимости концентрации вируса на поверхности и внутри клетки в зависимости от времени и начальной концентрации вируса и клеточных рецепторов позволяет получить кинетические параметры системы вирус гриппа A/Aichi/2/68 - рецепторы клеток MDCK, используя зависимость интенсивности флуоресценции от времени методом решения обратной задачи.
Используя данные, полученные методом иммуноферметного анализа, мы подобрали необходимые коэффициенты для корректной аппроксимации зависимости флуоресценции от времени. Расчеты проводились при подстановке в уравнение (3.12) начальной концентрации вируса [V0]= 3,5·10-13M и начальной концентрации свободных клеточных рецепторов [R0]= 3·10-10M.

По полученной аппроксимации сигнала флуоресценции, приведенной на рисунке 18, получена зависимость концентрации вируса участвующего в процессе эндоцитоза от времени.
Также по данным аппроксимации рассчитаны кинетические параметры системы вирус-клетка, находящейся при температуре 370C.
Таблица 2 Кинетические параметры системы «вирус - клетка», полученные методом флуоресцентной детекции
T, °C | k+, (М·min) -1 | k-,(М·min )-1 | kin,(М·min)-1 |
37 | 1,5·109 | 0,04 | 0,3 |
Приведенные в таблице 2 данные говорят о быстром осаждении вируса на клетку, так как максимальная концентрация вируса достигает [VR]m= 2·10-14М (Рисунок 18), а значит время насыщения клеточных рецепторов вирусом принимает значение tm= 2,6±0,5мин. (см. Теоретическая часть. Модель процесса эндоцитоза). Это можно объяснить не только самой оптимальной для данного процесса температурой (37°С), но и тем фактом, что и вирус и клетки находились в суспензии (в отличие от метода ИФА и ФОЕ, где клетки находились в монослое). Константа проникновения (kin) также достаточно высока, что объясняется быстрой адсорбцией вируса на клетку за счет насыщения системы «вирус - клетка» клеточными рецепторами.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
