
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
При наличии неизбежных сезонных, межгодовых и региональных различий при анализе осредненных данных нельзя не заметить и ряд общих и характерных черт. Так, повсеместно видна подчиненная роль в биомассах зоопланктона мелкой и средней фракций планктона. Доля крупной фракции, т. е. макропланктона, лишь за некоторыми исключениями укладывается в диапазон от 70 до 80-85 % от общей биомассы. Мелкая и средняя фракции в целом находятся примерно в равном соотношении. Однако нельзя не заметить, что доля средней фракции несколько увеличивается от весенне-летнего периода к осенне-зимнему, что связано с весенним размножением планктона и ростом ранних стадий планктеров. Если иметь в виду средние биомассы мелкой и средней фракций, то четких закономерностей при сравнении крупных районов не просматривается. Летом по концентрациям мелкого планктона районы могут быть перечислены в порядке: Берингово море, прикурильские воды океана, Охотское море, восточнокамчатские воды. Осенью этот порядок уже иной: Охотское море, Берингово море, прикурильские и прикамчатские воды. Наибольшая биомасса средней фракции и в летние и осенние сезоны наблюдается в Охотском море.
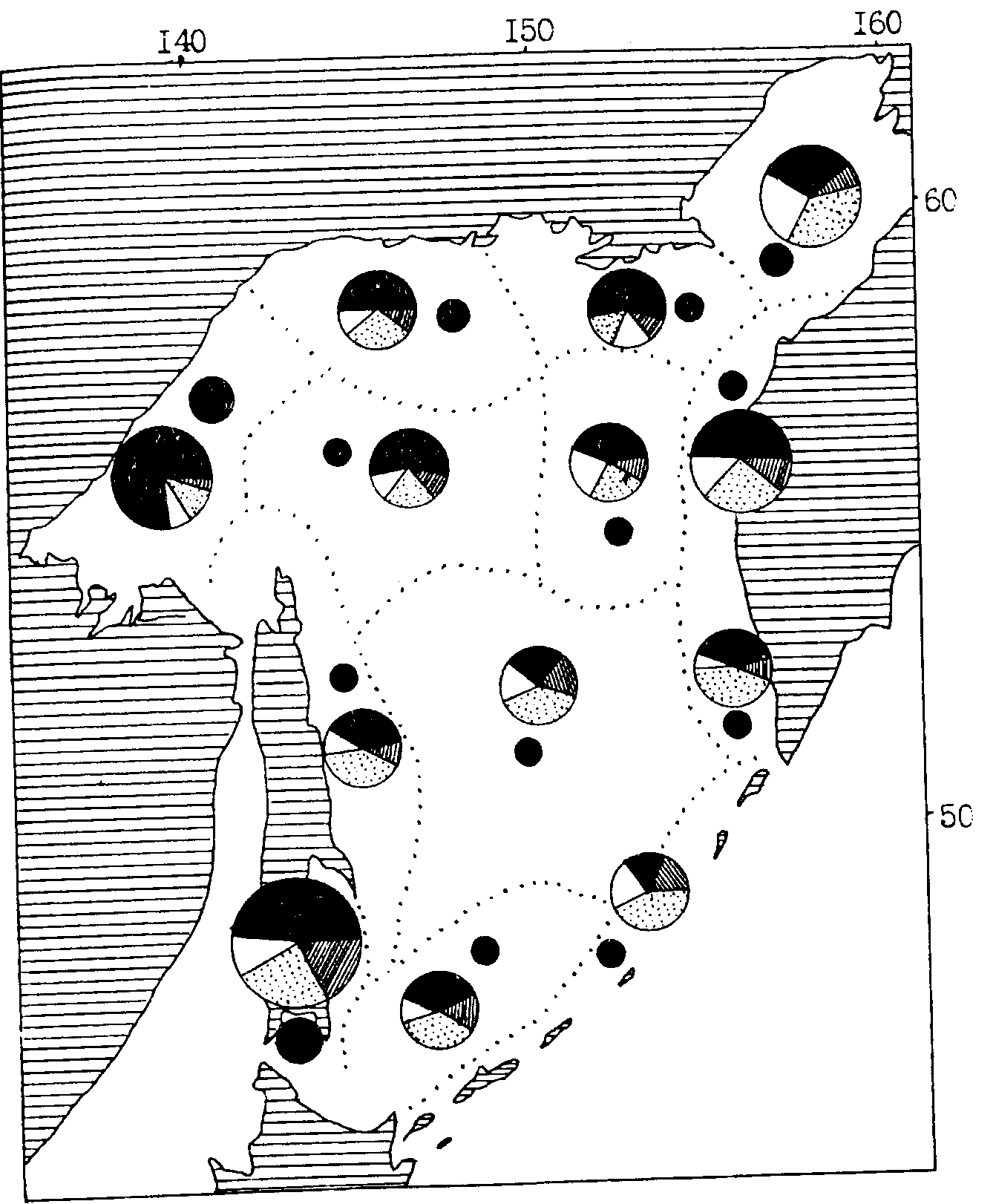
Рис. 101. Средняя биомасса зоопланктона в различных районах Охотского моря в осенний период (1984, 1985, 1987, 1990, 1991 гг). Обозначения как на рис. 100
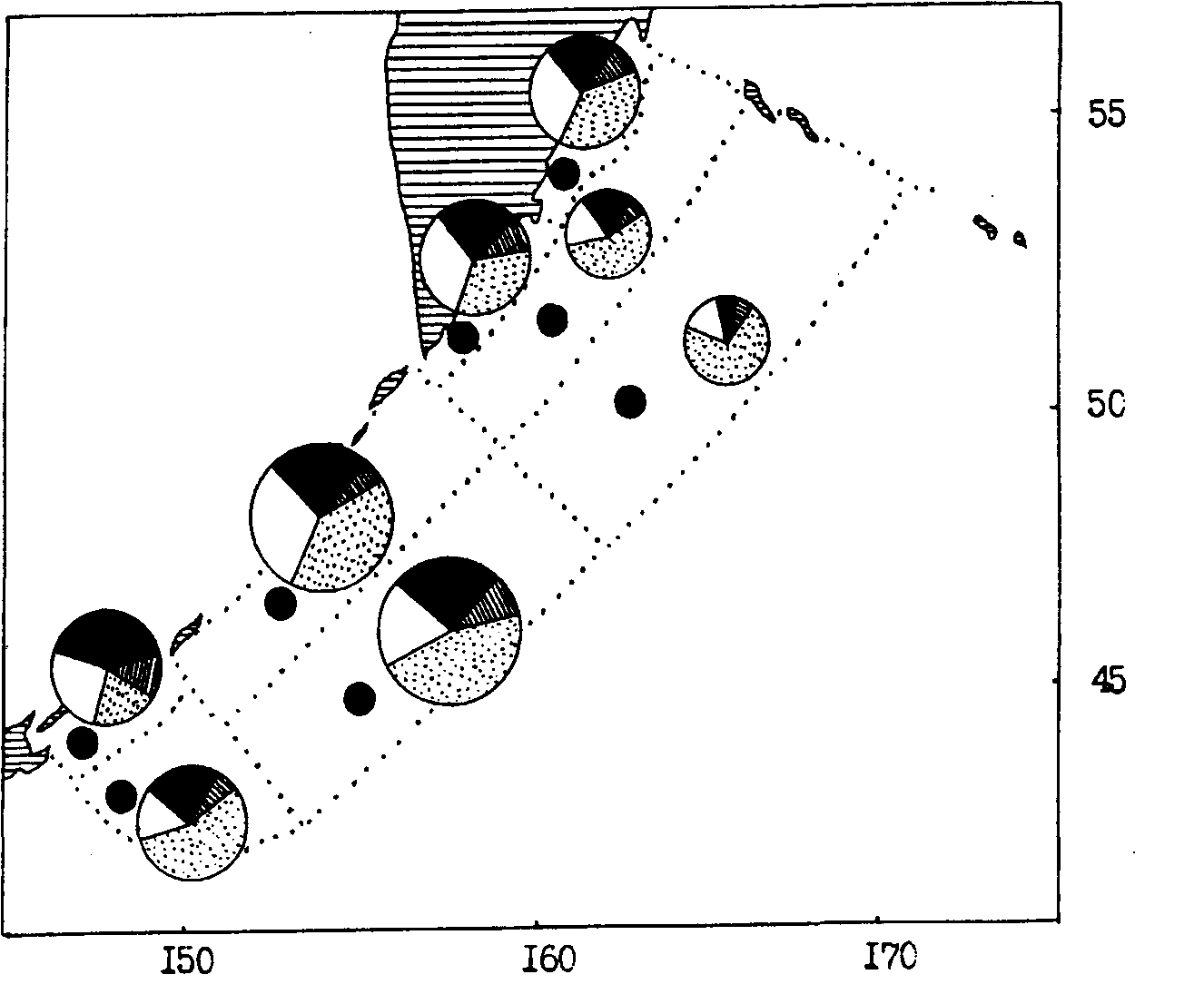
Рис. 102. Средняя биомасса зоопланктона в различных районах прикамчатских и прикурильских вод Тихого океана в летний период 1991 г. Обозначения как на рис. 100
Планктон мелкой и средней фракций, наряду с бактериями и простейшими, формирует кормовую базу хищного планктона и ранних стадий нектона. Основу биомассы мелкой фракции слагают Pseudocalanus minutus, Oithona similis, Acartia longiremis, яйца, личинки и науплии других планктеров (в первую очередь копепод и эвфаузиид) и донных беспозвоночных. В биомассах средней фракции доминируют все тот же Pseudocalanus minutus, копеподиты калянид, калиптописы и фурцилии эвфаузиид и личинки донных беспозвоночных.
Пока во многом не ясно, какое значение могут иметь указанные выше межрегиональные различия в биомассах мелкой и средней фракций для функционирования планктонных сообществ и кормовой обеспеченности ранних стадий нектона. Можно, однако, предположить, что более высокая биомасса мелкого планктона летом в Беринговом море должна позитивно сказываться на воспроизводстве некоторых видов нектона. Кстати, это обстоятельство, по-видимому, частично объясняет описанный выше факт, что беринговоморские мальки минтая несколько крупнее охотоморских.

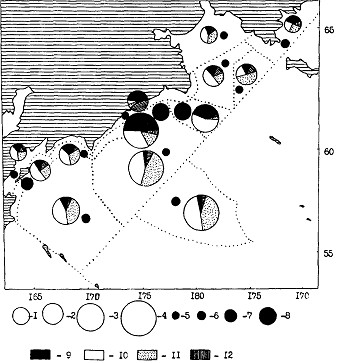
Рис. 103. Средняя биомасса зоопланктона в различных районах прикамчатских и прикурильских вод Тихого океана в осенний период (1986, 1987, 1991 гг.). Обозначения как на рис. 100
При анализе пространственного количественного распределения мелкого и среднего планктона по осредненным данным обращает на себя внимание довольно монотонный уровень средней биомассы в большинстве районов и подрайонов. Особенно это характерно для тихоокеанских вод Камчатки и Курильских островов (см. рис. 102, 103). В Охотском море (см. рис. 100, 101) повышенной биомассой мелкого и среднего планктона отличаются некоторые шельфовые районы и в первую очередь северо-восточная (районы 1, 2, 3, 7) и юго-западная (район 11) части моря. В Беринговом море (см. рис. 104, 105) в этом плане выделяются наваринские (районы 5 и 7) и карагинские (район 11) воды.
Вряд ли сильно отличается от описанной для более северных районов размерная структура планктонных сообществ в Японском море. Для северной части моря, по с соавторами (1990), на долю мелкого и среднего планктона приходится весной 55 %, летом 42-44, осенью 27, зимой 24 %, т. е. заметно больше, чем в Беринговом, Охотском морях с сопредельными водами океана (табл. 39-41). Но в данном случае из состава мелкой фракции не выделялся фитопланктон, что, несомненно, и определило завышенное в упомянутой работе количество мелкого планктона в Японском море. Особенно наглядно это видно на примере весеннего и летнего сезонов. Весной и в начале лета, т. е. во время цветения, плотность концентраций мелкой фракции (с фитопланктоном) составляет в разных районах Японского моря соответственно около 38-80 и 31-52 г/м, т. е. выше, чем в северных водах. Во второй половине лета, когда цветение в основном заканчивается, плотность концентраций мелкой фракции уменьшается до 12-18 г/м (Лапшина и др., 1990), что соответствует аналогичным
данным в Беринговом и Охотском морях (табл. 39-41).
Рис. 104. Средняя биомасса зоопланктона в различных районах западной части Берингова моря в летний период (1989, 1991 гг.). Макропланктон: 1- менее 50 г/м, 2 -50-100, 3 - 100-150, 4 - более 150; мелкий и средний планктон: 5 - менее 50, б - 50-75, 7-75-100, 8- более 100 г/м2; 9 - эвфаузииды, 10 - копеподы, 11- сагитты, 12 - прочие
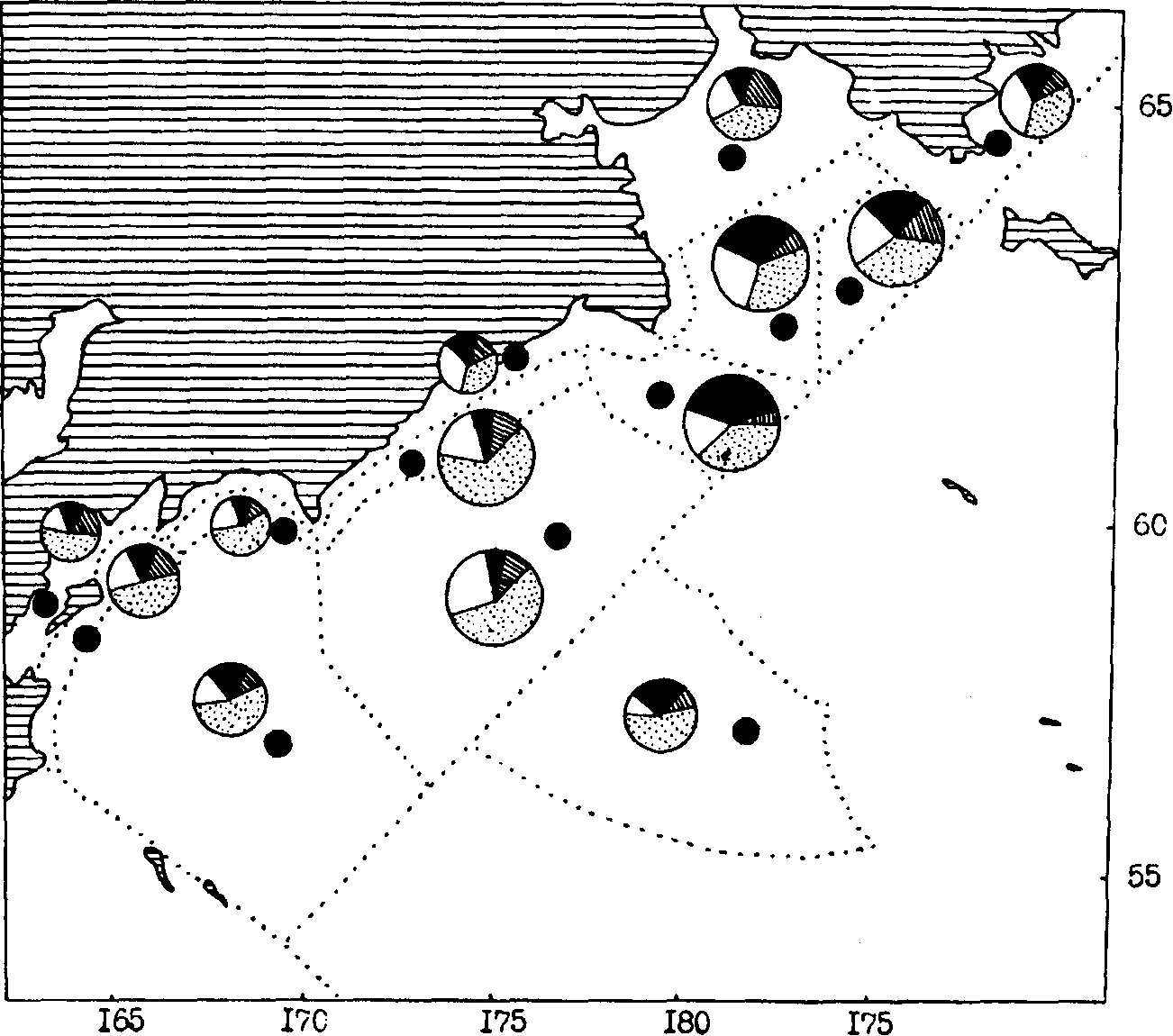
Рис. 105. Средняя биомасса зоопланктона в различных районах западной части Берингова моря в осенний период (1986, 1987, 1988, 1990гг.). Обозначения как на рис. 104
Крупную фракцию формирует довольно большая группа планктеров - эвфаузиид (в основном Thysanoessa raschii, T. longipes, T. inermis), копеподы (в основном Calanus plumchrus, C. cristatus, C. glacialis, Eucalanus bungii, Metridia okhotensis, M. pacifica, Bradyidius pacificus), амфиподы (в основном Parathemisto pacifica, P. libellula) и, конечно, сагитты (в основном Parasagitta elegans).
Видовой состав доминирующих видов в каждом конкретном случае зависит от биотопического фона и положения рассматриваемого участка в системе биогеографических подразделений.
Количественное распределение макропланктона более контрастно по сравнению с распределением более мелкого планктона (табл. 39-41, рис. 100-105). В летний период наиболее высокой биомассой крупной фракции выделяются прикурильские воды Тихого океана, что, несомненно, находится в связи с очень высокой и сложной динамикой вод этого района. Однако осенью концентрации планктона в верхнем 200-метровом слое здесь находятся на уровне величин смежных районов. Это выравнивание, судя по всему, связано со значительным выеданием продукции мирного планктона, а также большим размахом сезонных вертикальных миграций в условиях больших глубин данного района.
Во все сезоны концентрации макропланктона в Охотском море гораздо выше, чем в Беринговом и смежных с ним тихоокеанских водах Камчатки. На это обстоятельство выше уже обращалось внимание в связи с обсуждением питания минтая и его кормовой обеспеченностью.
К сожалению, нет полностью сопоставимых данных по плотности концентраций макропланктона в Японском море. В работе с соавторами (1990), где суммируются результаты многих планктонных съемок в северной части Японского моря в 80-е гг., не вводились поправки, учитывающие суточные вертикальные миграции макропланктона, к тому же облавливался только верхний 100-метровый слой. Все это не могло не привести к занижению концентраций планктона. По данным упомянутых авторов, биомасса макропланктона в северной части Японского моря составляет весной 45-72, в первой половине лета - около 83-140, во второй половине лета - 21-51, осенью - 63, зимой -31 г/м. Только данные за первую половину лета соответствуют аналогичным показателям в Беринговом и Охотском морях, во всех остальных случаях они ниже в 1,5-2,0 раза (табл. 39-41). Учитывая замечания, относящиеся к методике расчетов плотности концентраций планктона в Японском море, можно, по-видимому, сделать предположение о том, что они приближаются к значениям, характерным для Берингова моря и тихоокеанских вод Камчатки.
Весьма существенным моментом с точки зрения оценки кормовых условий нектона в различных районах является состав макропланктона и его динамика. Например, доля эвфаузиид, являющихся одной из наиболее предпочитаемых групп кормовых объектов нектона, во все сезоны наиболее высокой бывает в эпипелагиали Охотского моря (см. табл. 40, рис. 100,101). По биомассе обычно они занимают первое место, лишь летом уступая копеподам. Вторыми эвфаузииды идут и в прикурильских водах океана, но здесь, а также в восточнокамчатских водах количественно даже летом преобладают сагитты. В Беринговом море на 1-2-х местах обычно стоят копеподы.
При сопоставлении данных табл. 39-41 хорошо видно, что в большинстве крупных районов в эпипелагиали от лета к осенне-зимнему периоду уменьшается доля копепод, но вместе с тем увеличивается дол я эвфаузиид, амфипод и сагитт. Эта динамика является итогом наложения по крайней мере трех процессов. Здесь можно уверенно говорить, например, о влиянии неодинаковых у разных групп амплитуд сезонных вертикальных миграций, а также темпа сезонного роста. Наконец, это связано с выеданием мирного планктона хищным, что, как известно, и образует один из важнейших механизмов сезонных сукцессии в сообществах, в результате чего осенью количество первых уменьшается, а вторых возрастает.
О выедании планктона ниже пойдет специальный разговор, здесь же кратко коснемся динамики вертикального распределения планктона. Вся приведенная выше информация относится только к эпипелагиали. На шельфе при обловах слоя 0-200 м в принципе весь планктон должен находиться в зоне действия сети в любое время суток. Однако и здесь дневные уловы, как правило, заметно ниже. Это объясняется концентрациями части планктеров непосредственно в придонных слоях, где их облов затруднен технически. Что касается больших глубин, то в этом случае значительная часть планктеров или уходит в светлое время суток из верхних слоев или вообще не поднимается в эпипелагиаль. При этом следует оговориться, что крупномасштабные миграции совершает только макропланктон (табл. 42).
Таблица 42
Изменение биомассы зоопланктона на разных глубинах и течение суток и южной част и Охотского моря в январе 1992 г. (суточная станция), мг/м3

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 |
