
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
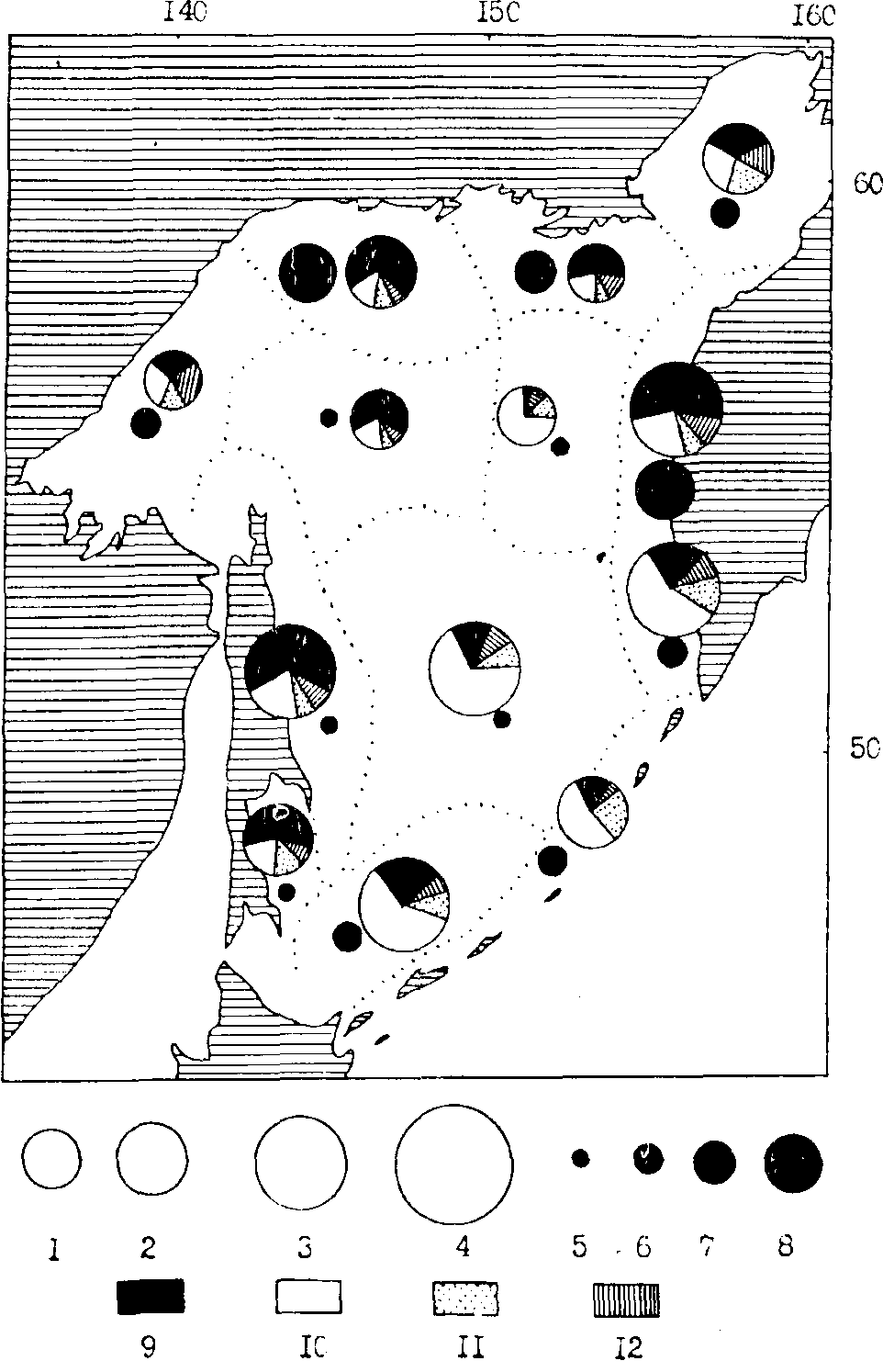
Рис. 106. Средняя биомасса зоопланктона в различных районах Охотского моря в июле-сентябре 1986 г. Макронланктон: 1 - менее 1000 мг/м3, 2 - 1000-1500, 3 - 1500-2000, 4 - более 2000; мелкий и средний планктон: 5 - менее 250,6 - 250-500, 7 - 500-1000, 8 - более 1000 мг/м, 9 - эвфаузииды, 10 - копеподы, 11 - сагитты, 12 - прочие
Указанные и другие межгодовые различия являются результатом наложения и суммирования биомасс конкретных, в первую очередь массовых, видов планктеров, каждый из которых имеет свою динамику численности. К сожалению, на данный момент изученность экологии, в том числе жизненных циклов и экологических потребностей абсолютного большинства видов планктеров дальневосточных морей, находится в основном на начальных стадиях. В связи с этим даже предположительно трудно или невозможно назвать конкретные факторы, благоприятствующие или препятствующие воспроизводству того или иного вида. Хорошо же известных общих представлений о важности для количественного развития планктона таких факторов, как температурный режим, интенсивность вертикальной и горизонтальной динамики вод, количество фитопланктона и т. д., здесь явно недостаточно. Отсюда неизбежны пока и просто констатация фактов и большие допущения, а также применение аналогов из других более изученных районов. Неизбежно, как и ранее, оперирование осредненными биомассами зоопланктона в целом, тем более что многие массовые рыбы демонстрируют большую пластичность в питании и свободно переходят с одного кормового объекта на другой.
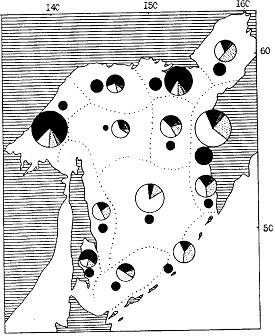
Рис. 107. Средняя биомасса зоопланктона в различных районах Охотского моря в июне-августе 1988 г. Обозначения как на рис. 106
Межгодовые изменения в обилии планктона и его количественном распределении не могут не отражаться на распределении нектона и, конечно, такого массового вида как минтай. Тем не менее при элементарных сопоставлениях биомасс рыб и планктона в большинстве случаев не удается заметить определенных закономерностей. По крайней мере ни одна из многочисленных экспедиций последних лет, по материалам которых выше описано питание минтая и даны некоторые количественные характеристики планктонных сообществ, не подтвердила широко распространенные схемы типа "много биогенов - много планктона - много рыбы" или "мало биогенов - мало планктона — мало рыбы" (Шунтов и др., 1988а, б, 1990а; Sobolevskiy et al., 1989; Ефимкин, Радченко, 1991). В этом смысле весьма характерны ситуации, показанные на рис. 110-112. При их анализе нужно иметь в виду, что в большинстве районов среди рыб преобладает минтай.
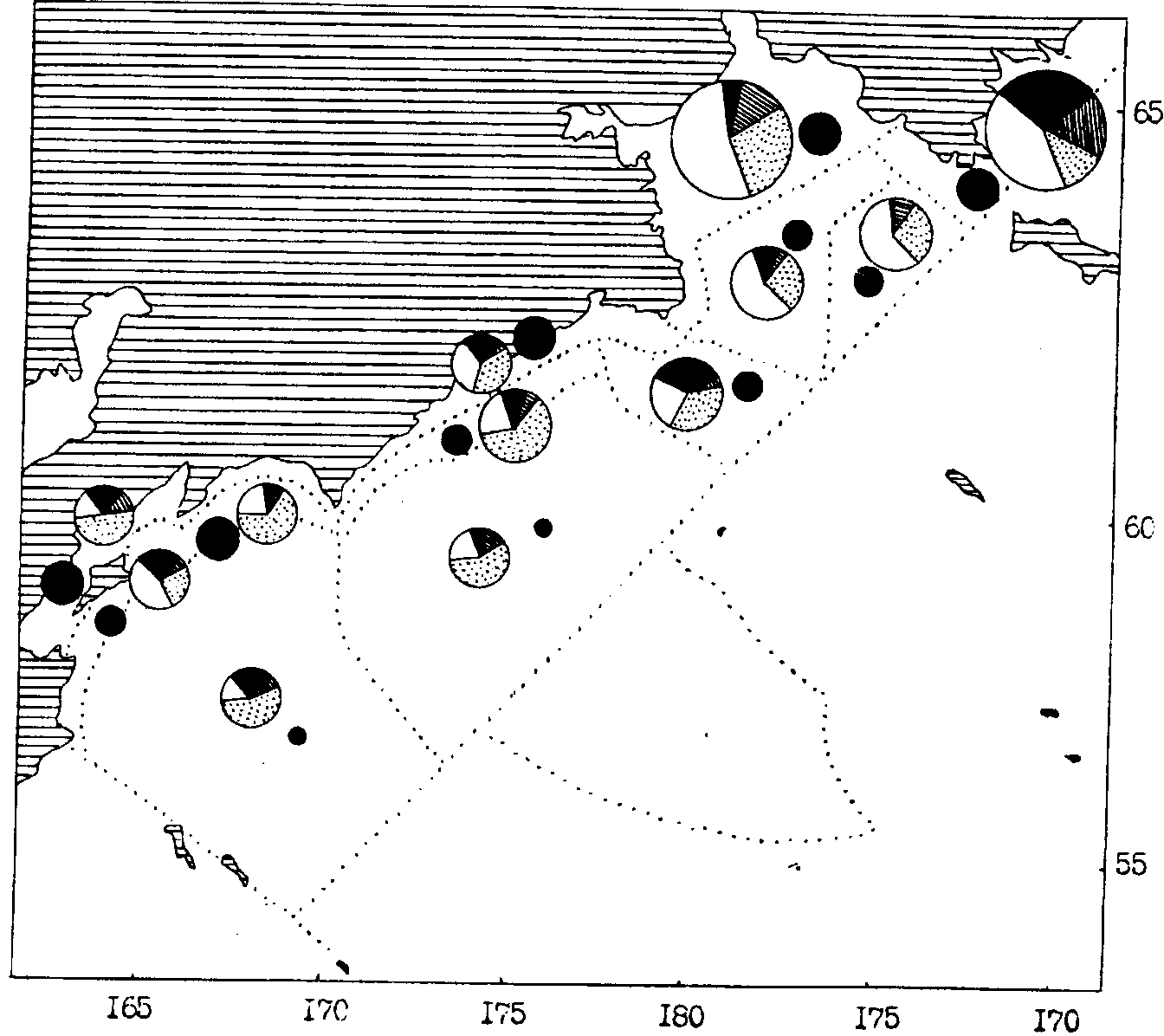
Рис. 108. Средняя биомасса зоопланктона в различных районах западной части Берингова моря в сентябре-октябре 1986 г. Обозначения как на рис. 106
Обращая внимание на отсутствие простых связей в системе "планктон - нектон", мы не имеем в виду вообще ставить под сомнение известные общие закономерности формирования продуктивных зон и их влияние на численность и распределение нектона. В разделе о питании минтая уже приводились некоторые фактические данные в пользу выводов о важности для него не только обилия и состава кормовых объектов, но и других показателей, определяющих структуру кормовой базы. Одним из важных моментов здесь, по-видимому, является характер формирования кормовых полей и слоев с более высокими концентрациями кормовых организмов. Для более определенных суждений в таких вопросах конкретных данных пока мало, особенно информации о тонкой структуре океанологических полей (мезо - и микрозавихрения, вторичные фронты и т. д.)., переносе и накоплении планктона в конкретных локальных районах. Последнее обстоятельство, судя по всему, вообще может играть в ряде случаев решающую роль в формировании скоплений планктона, а следовательно, и нектона. В первую очередь в этом плане необходимо назвать некоторые участки в зоне материкового склона: в Охотском море - западнокамчатский район и склоны впадины ТИНРО, а в Беринговом - наваринско-карагинский район. На этих участках в течение всего года держатся значительные концентрации неполовозрелого минтая, а временами и половозрелых особей. В то же время здесь далеко не всегда бывают высокими биомассы зоопланктона. Тем не менее скопления рыб не рассеиваются и не покидают эти участки. Следовательно, планктон здесь должен постоянно пополняться за счет привноса со смежных акваторий. В зоне свала глубин процессу формирования кормовых полей несомненно способствуют фронтальные образования, которые благодаря орографическому эффекту должны занимать относительно стабильное положение. Кроме того, в водах материкового склона в связи с высокой динамикой вод создаются благоприятные условия для развития и местного планктона.
В основном иная картина в смысле стабильности должна наблюдаться в глубоководных котловинах и на шельфах, где океанологические границы более размыты или подвижны. В такой ситуации и скопления рыб должны быть более подвижными, что и наблюдается в действительности. Исключением могут быть только четкие океанологические разделы типа притауйского фронта или фронта по южной границе анадырско-матвеевского холодного пятна.
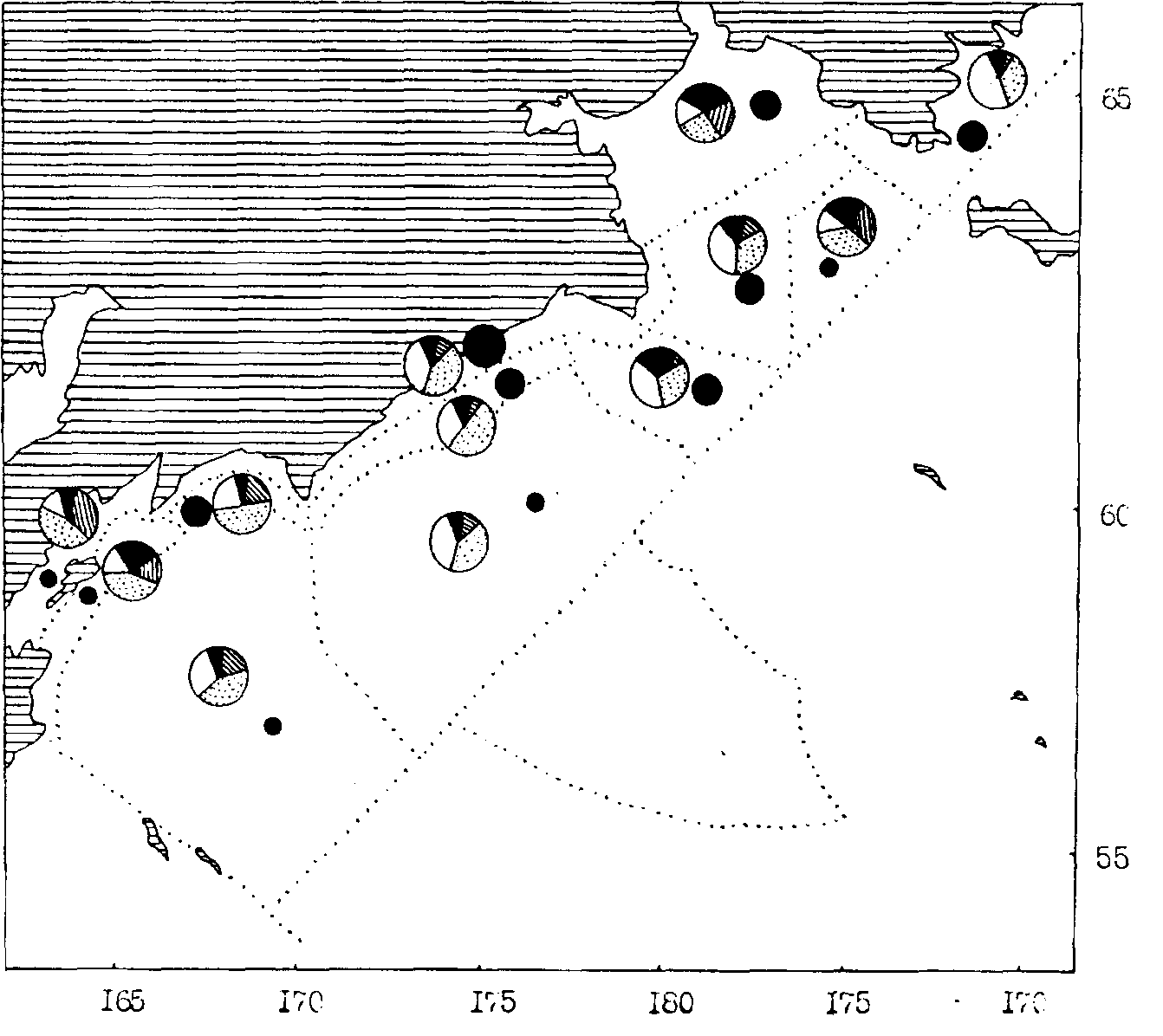
Рис. 109. Средняя биомасса зоопланктона в различных районах западной части Берингова моря в сентябре-октябре 1987 г. Обозначения как на рис. 106
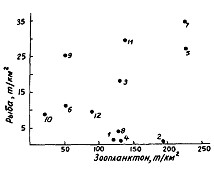
Рис. 110. Соотношение плотности концентраций рыб и зоопланктона крупной и средней фракции в сентябре—октябре 1986 г. в Беринговом море (Шунтов и др., 1988а): 1-12 - районы как на рис. 62
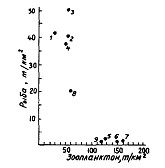
Рис. 111. Соотношение плотности концентраций рыб и зоопланктона крупной и средней фракции в ноябре-декабре 1986 г. в тихоокеанских водах Камчатки и Курильских островов (Шунтов и др., 19886): 1-9 - районы как на рис. 60

Рис. 112. Соотношение плотности концентраций макропланктона и рыб В эпипелагиали различных районов Охотского моря в июне-августе 1988 г. (Шунтов и др., 1990а): 1 -13 - районы как на рис. 61
Изложенное выше в некоторой степени носит предположительный характер, так как проблема формирования планктонных кормовых поле, а точнее его механизмы, слабо изучены, а применительно к рассматриваемым нами районами вообще не известна. В связи с этим заметим, что пятнистость (скопления, агрегации, стаи) в распределение планктона не только может зависеть от гидродинамических факторов, но также формируется на базе поведенческих реакций и трофических отношений (Зеликман, 1977) В упомянутом здесь обзоре, например, описывается здесь скопления ойкоплевры шириной в несколько метров и длиной в несколько километров, возникающие в районе цветения воды фитопланктоном. Образования этих агрегаций связываются с действием мелкомасштабных горизонтальных циркуляций, возникающих на линиях конвергенций. Ясно, что с энергетической точки зрения целенаправленное питание рыб в таких случаях бывает эффективным. Кстати, при описании питания минтая в северо-западнокамчатском районе похоже ситуация с переключением его на ойкоплевру уже упоминалась.
Более изучена пятнистость распределения ракообразных и в первую очередь эвфаузиид. Рыбы, несомненно, обладают необходимым набором поведенческих адаптаций для поиска таких скоплений, при этом в качестве ориентиров вполне могут использовать различные гидродинамические и физико-химические факторы.
Для нектона, помимо плотности концентраций и наличия скоплений планктона, важное значение должны иметь объем общих планктонных ресурсов в том или ином районе или в море в целом. При больших запасах планктона, даже если они рассредоточены на обширных площадях или в больших объемах вод, по-видимому, есть большие основания ожидать формирование кормовых поле при наличии определенных гидрологических ситуаций. Показательны в этом смысле некоторые соотношения, полученные при сравнении различных районов Охотского и Берингова морей и сопредельных с ними вод Тихого океана.
В эпипелагиали Охотского моря (таб. 44) летом примерно половины макропланктона (49-55%) и всего зоопланктона (48-51,6%) сосредоточена в глубоководной котловине (без впадин ТИНРО и Дерюгина). Осенью в связи со смещением продуктивных вод на север, а также уходом части планктеров из эпипелагиали в более низкие горизонты, доля биомассы в глубоководной котловине несколько уменьшается (макропланктон - 31,1-47,1%, а весь зоопланктон - 31,8-45,7%). Но и в этом случае эта доля измеряется цифрами, близкими к 100 млн. т и половины биомассы планктона сосредоточенно ниже 200 м, можно видеть исключительную масштабность планктонных ресурсов этой части моря.
Таблица 44
Общий запас зоопланктона и эпинелагиали различных районов Охотского моря
Объект | Шельф и материковый склон, районы 1-4, 7, 8, 10, 11 | Впадины ТИНРО и Дерюгина, районы 5, 6 | Глубоководная часть, районы 9, 12, 13 | Всего |
Июль-сентябрь 1989 г. | ||||
Макропланктон млн. т | 76,5 | 39,5 | 142,1 | 258,1 |
% | 29,7 | 15,3 | 55,0 | 100,0 |
Весь зоопланктон млн. т | 107,3 | 48,0 | 165,6 | 320,9 |
% | 33,4 | 15,0 | 51,6 | 100,0 |
Июнь-август 1988 г. | ||||
Макропланктон млн. т | 110,5 | 47,2 | 151,5 | 309,2 |
% | 35,7 | 15,3 | 49,0 | 100,0 |
Весь зоопланктон млн. т | 153,0 | 60,5 | 196,7 | 410,2 |
% | 37,3 | 14,7 | 48,0 | 100,0 |
Октябрь-ноябрь 1984 г. | ||||
Макропланктон млн. т | 88,7 | 30,0 | 53,4 | 172,1 |
% | 51,5 | 17,4 | 31,1 | 100,0 |
Весь зоопланктон млн. т | 145,4 | 37,9 | 85,5 | 268,8 |
% | 54,1 | 14,1 | 31,8 | 100,0 |
Сентябрь-ноябрь 1985 г. | ||||
Макропланктон млн. т | 65,5 | 51,7 | 104,3 | 221,5 |
% | 29,6 | 23,3 | 47,1 | 100,0 |
Весь зоопланктон млн. т | 96,0 | 74,1 | 143,2 | 313,3 |
% | 30,6 | 23,7 | 45,7 | 100,0 |
Октябрь 1987 г. | ||||
Макропланктон млн. т | 85,9 | 42,7 | 72,7 | 201,3 |
% | 42,7 | 21,2 | 36,1 | 100,0 |
Весь зоопланктон млн. т | 122,7 | 67,3 | 104,3 | 294,3 |
% | 41,7 | 22,9 | 35,4 | 100,0 |
Площадь, тыс. км2 | 630 | 280 | 592 | 1502 |
Примечание. Районы как на рис. 61.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 |
