

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Через 48 ч после введения ЛПС у мышей контрольной группы отмечали прогредиентное увеличение относительной массы селезёнки, доли белой пульпы и уменьшение ширины маргинальной зоны (табл. 7). Количество клеток в маргинальной зоне и красной пульпе не претерпевало количественных изменений, но наблюдали значительное снижение содержания нейтрофилов (табл. 7). Тучные клетки не были выявлены (табл. 7). Расширения кровеносных сосудов красной пульпы не наблюдали.
Через 48 ч после введения ЛПС масса селезёнки мышей опытной группы уменьшена в отличие от контрольной группы и не отличалась от значений до введения ЛПС (табл. 7). Доля белой пульпы меньше, чем в контрольной группе (табл. 7). Ширина маргинальной зоны уменьшена и не отличалась от контрольной группы. Количество клеток в маргинальной зоне также уменьшено, и было немногим больше, чем в контрольной группе, нейтрофилы практически не выявляли. Наблюдали прогрессирующее опустошение красной пульпы. В отличие от контрольной группы в ней выявляли крайне низкое число нейтрофилов, но встречали единичные тучные клетки с пониженным содержанием секреторных гранул (табл. 7).
Таким образом, стимуляция иммунной системы материнского организма в ранние сроки беременности приводит к менее выраженной реакции тимуса и селезёнки потомства при развитии системного воспалительного ответа, что связано с функциональной незрелостью иммунной системы.
Табл. 7
Морфофункциональные характеристики селезёнки мышей контрольной и опытной групп в разные сроки после введения ЛПС (M±m)
Сроки исследования Морфологические показатели | До введения ЛПС | Через 24ч после введения ЛПС | Через 48ч после введения ЛПС |
Относительная масса селезёнки, % Контрольная группа Опытная группа | 0,64±0,06 0,57±0,04 | 0,72±0,06# 0,71±0,06# | 0,97±0,08♦ 0,64±0,05* |
Доля белой пульпы, % Контрольная группа Опытная группа | 26,59±4,66 19,62±2,75* | 30,74±0,04# 26,61±1,28*# | 42,38±3,25♦ 30,45±2,33* |
Ширина маргинальной зоны, мкм Контрольная группа Опытная группа | 60,72±3,51 45,55±6,35* | 36,76±2,27# 34,63±1,73# | 25,0±1,75♦ 21,97±1,91♦ |
Количество клеток в мм2 среза маргинальной зоны Контрольная группа Опытная группа | 21375,00±781,20 29652,50±868,90* | 9922,50±1256,50# 19468,30±1522,00*# | 9290,00±461,50 11802,50±787,80#♦ |
% нейтрофилов от общего количества клеток в маргинальной зоне Контрольная группа Опытная группа | 1,04±0,05 0,13±0,01* | 20,15±2,57# 0,63±0,07*# | 3,41±0,31♦ 0,21±0,01*#♦ |
Количество клеток в мм2 красной пульпы Контрольная группа Опытная группа | 24175,00±803,30 27275,00±1076,20* | 11391,40±450,40# 18144,40±491,00*# | 11252,90±1224,90 14362,50±602,10*♦ |
% нейтрофилов от общего количества клеток в красной пульпе Контрольная группа Опытная группа | 0,27±0,02 0,09±0,005* | 6,28±0,89# 0,98±0,11*# | 2,65±0,54♦ 0,21±0,09*♦ |
Количество тучных клеток в мм2 площади среза селезёнки Контрольная группа Опытная группа | 0,92±0,31 5,49±0,25* | 0,49±0,15 0,55±0,11# | 0 1,02±0,36* |
СГК тучных клеток Контрольная группа Опытная группа | 0,75±0,04 1,46±0,06* | 2,23±0,23# 2,67±0,33# | – 1,88±0,12♦ |
ИД тучных клеток Контрольная группа Опытная группа | 0 0 | 0 33,33±2,50*# | – 0♦ |
Примечания: * - статистически значимые различия между опытной и контрольной группами; # - между значениями до введения ЛПС и через 24ч после введения ЛПС; ♦ - между значениями через 24ч после введения ЛПС и через 48ч после введения ЛПС.
Реагирование иммунной системы потомства самок мышей, перенесших однократную стимуляцию иммунной системы в ранние сроки беременности, при развитии опухолевого процесса
При исследовании цитотоксической активности супернатантов, полученных при культивировании клеток селезёнки с клетками опухоли, обнаруживали дозозависимый лизис клеток L929 как в опытной, так и в контрольной группах. Однако, процент цитолиза в контрольной группе животных был статистически значимо выше, чем у мышей рождённых от самок, получавших Кон А (рис. 3).
Исследование развития опухолевого процесса у мышей контрольной группы и потомства самок, перенесших однократную стимуляцию иммунной системы на ранних сроках беременности
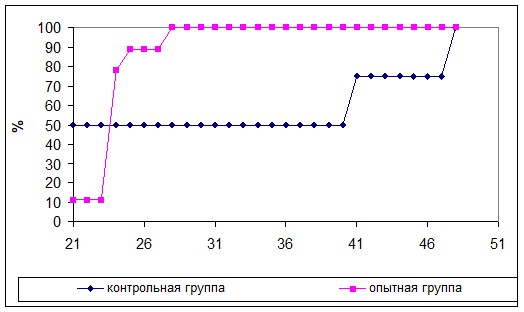
При ежедневном исследовании животных, которым подкожно вводили клетки меланомы В16, на 21-ые сутки после введения опухолевых клеток пальпируемая опухоль обнаруживали у 18,2% животных контрольной группы и 8,3% животных опытной группы. В дальнейшем в опытной группе наблюдали резкое увеличение приживаемости опухоли в отличие от контрольной группы. На 28-ые сутки этот показатель достиг максимального значения – 75%. В контрольной группе животных опухоль привилась у 36,4% мышей.
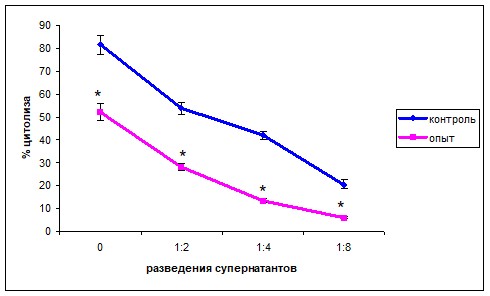
Рис. 3. Цитотоксическая активность клеток селезёнки мышей, родившихся от самок, получавших КонА, и мышей контрольной группы в отношении клеток фибросаркомы L929 (M±m)
Примечание: * - статистически значимые отличия от значений контрольной группы, 0 по оси абсцисс – без разведения.
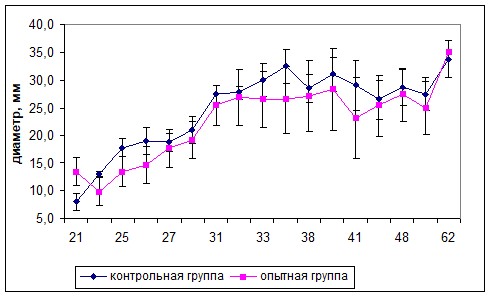
Исследования темпов приживаемости среди мышей, у которых она привилась, выявляли более высокие значения этого показателя у мышей опытной группы (рис. 4а). Размеры опухоли у животных обеих групп статистически значимо не отличались в течение исследуемого периода вплоть до гибели животных (рис. 4б).
a б
Рис. 4. Приживаемость меланомы В16 (а) и темпы роста подкожной опухоли меланомы В16 (M±m) (б) у мышей опытной и контрольной групп
Темпы гибели животных-опухоленосителей опытной группы были выше. Так гибель 50% мышей в опытной группе наступила на 7 сут. раньше, а гибель 75% – 17 сут. раньше, чем в контрольной группе (рис. 5). При вскрытии погибших животных во всех случаях обнаруживали прорастание опухоли в органы брюшной полости.
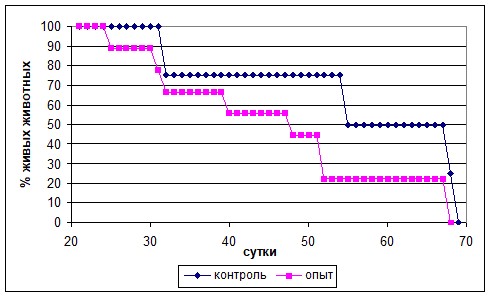
Рис. 5. Продолжительность жизни мышей-опухоленосителей, родившихся от самок, получавших Кон А, и контрольной групп
Таким образом, стимуляция иммунной системы материнского организма на ранних сроках беременности приводит к ослаблению противоопухолевого иммунитета у потомства вплоть до периода полового созревания. Одним из механизмов снижения противоопухолевого иммунитета является уменьшение цитотоксической активности лимфоцитов, уменьшение количества NK - и NKТ-клеток и снижение способности синтезировать ФНО-б в ответ на опухолевые антигены.
Морфофункциональные изменения кожи и её производных потомства самок мышей, перенесших однократную стимуляцию иммунной системы в ранние сроки беременности, в разные периоды постнатального морфогенеза
На 17-ые сутки постнатального развития, когда происходит первое цикличное изменение роста волос у мышей [Muller-Rover S. et al., 2001; Fuchs E., 2007] у потомства самок, перенесших иммуностимулирующее воздействие, развивалась транзиторная очаговая алопеция. Утрата шерстного покрова, начиналась со спины и распространялась на брюхо, но не затрагивала кожу головы, лап и основания хвоста (рис. 6). Большинство волос вокруг очага алопеции находились в стадии телогена. При исследовании гистологических препаратов участка кожи спины, находящегося рядом с очагом выпадения волос и сохранившем шерстный покров, выявляли утолщение мальпигиева слоя эпидермиса (табл. 8), очаговое снижение интенсивности ШИК-реакции в базальной мембране, размытость базальной мембраны. Толщина дермы практически не отличалась от значений контрольной группы (табл. 8). Отмечали уменьшение количества аморфного вещества межклеточного матрикса по всей толщине дермы, благодаря чему она имела рыхлое сетчатое строение, волосяных фолликулов и сальных желёз (табл. 8). Выявлены уменьшение числа волосяных фолликулов и сальных желёз. Большинство волосяных фолликулов содержало терминальные волосы, количество пуховых волос было уменьшено. Многие волосяные фолликулы были деформированы, имели кистообразный вид и содержали неправильно ориентированные, изогнутые волосы с неравномерным распределением пигмента в стержне волоса. У многих волосяных фолликулов дермальное корневое влагалище характеризовалось слабо положительной ШИК-реакцией. Общее количество клеток в дерме было меньше (табл. 12). Также было меньшим число выявляемых тучных клеток (табл. 9). Большинство их отличалось высокой насыщенностью секреторным материалом. В целом, интенсивность выделения секреторных гранул за пределы клеток была ниже, чем в контрольной группе. Изменена и локализация тучных клеток в дерме и гиподерме. Многие тучные клетки располагались вдоль волосяных фолликулов. Изменений сосудистого русла не наблюдали.
При макроскопическом исследовании участка кожи с утраченным шерстным покровом отмечали сохранение единичных терминальных волос и отсутствие пуховых волос. Кожа имела бледно-розовую окраску. При микроскопическом исследовании кожа имела те же особенности строения эпидермиса и дермы, что и в пограничной зоне. В дерме выявляли большее число нейтрофилов, которое не наблюдали ни в пограничной зоне, ни в контрольной группе. Количество тучных клеток не отличалось от значений в контрольной группе. Их насыщенность секреторным материалом была выше, а интенсивность дегрануляции ниже (табл. 9). Сохранялась связь тучных клеток с волосяными фолликулами. Таким образом, окружение тучными клетками волосяных фолликулов и выделение ими секреторных продуктов предшествовало выпадению волос, что согласуется с данными о роли тучных клеток в развитии алопеции [Xu X. et al., 2003; Won C. et al., 2008; Vandrford D. et al., 2010]. В течение последующих 2-3 суток отмечали прогрессирующую утрату волосяного покрова спины и брюха. Волосяной покров сохранялся на голове, лапах и у основания хвоста. При гистологическом исследовании кожи головы наблюдали топографическое изменение структуры дермы. Фронтально расположенный участок кожи головы имел строение дермы аналогичное дерме животных контрольной группы. По направлению к цервикальному отделу в дерме содержание аморфного компонента межклеточного матрикса уменьшалось, но было больше, чем в коже спины. В коже спины отмечали пониженное содержание клеток по сравнению с контрольной группой и уменьшение числа нейтрофилов. Волосяные фолликулы были резко увеличены в размерах, имели кистообразное строение и не имели содержимого или содержали деформированные волосы. Всё вышесказанное позволяет сделать вывод, что выпадение и нарушение роста новых волос является результатом аномального морфогенеза как самих волосяных фолликулов и волос, так и дермы.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 |
