

72Трансляция – процесс построения полипептидной цепи по иРНК. Трансляция у всех организмов (прокариот и эукариот) происходит в цитоплазме на рибосомах при участии ферментов, тРНК и аминокислот, с затратой энергии (АТФ). Трансляция подразделяется на инициацию, элонгацию, терминацию. Инициация трансляции представляет собой событие, в ходе которого происходит образованиe комплекса, включающего мРНК и рибосому. Первоначально рибосомы соединяется с фактором инициации. Затем этот теперь уже более сложный комплекс связывается с концом мРНК. Один из факторов инициации присоединяется к участку на молекуле мРНК. Прикрепившись к мРНК начинает скользить по некодирующей части мРНК до тех пор, пока не достигнет инициирующего кодона кодирующей нуклеотидной последовательности. Скольжение по мРНК сопровождается гидролизом АТФ, энергия которого затрачивается на преодоление участков спирализации в нетранслируемой части мРНК. Достигнув начала кодирующей последовательности мРНК, происходит связывание с другими факторами инициации за счёт гидролиза ГТФ до ГДФ и неорганического фосфата. При этом формируются новые центры рибосомы, Элонгация-это наращивание (удлинение) пептидной цепочки, как и весь процесс трансляции, происходит на рибосомах.
73Терминация трансляции наступает в том случае, когда в центр рибосомы попадает один из стоп-кодонов. Для стоп-кодонов нет соответствующих тРНК. Вместо этого к рибосоме присоединяются 2 белковых фактора терминации. Один из них с помощью пептидилтрансферазного центра катализирует гидролитическое отщепление синтезированного пептида от тРНК. Другой за счёт энергии гидролиза ГТФ вызывает диссоциацию рибосомы. В целом же рост и функционирование митохондрии невозможны без импорта белков. Каждый поступивший белок должен достичь определенного субкомпартмента, в котором он функционирует. И каждый из этих субкомпартментов содержит отличный от других набор белков. Рост митохондрий возможен за счет импорта цитоплазматических белков, включающего последовательный избирательный перенос белков через одну или две мембраны. В большинстве случаев энергия, затрачиваемая на перемещение, используется в виде АТФ.
74Все гены находятся в большой самовоспроизводящейся молекуле ДНК. Каждый из них представляет собой небольшой участок такой молекулы. Но по своим функциям гены неодинаковы. Одни из них несут информацию о последовательности аминокислот в. белковой молекуле, т. е. определяют ее структуру, другие регулируют активность первых и контролируют тем самым процесс поступления информации от ДНК к и-РНК. Первая группа генов, получила название Структурных, Вторая—Регуляторных. Структурные гены, контролирующие синтез ферментов в какой-то одной цепи реакций, расположены обычно рядом друг с другом. Они составляют единый блок, и осуществляют последовательные этапы синтеза одного фермента, работая согласованно, как один элемент. Структурный ген - любой ген, кодирующий какую-либо полипептидную цепь или молекулу РНК, включая регуляторные гены, которые кодируют продукты, определяющие другие структурные гены. РНК-полимераза - Фермент, осуществляющий матричный синтез РНК. В зависимости от используемой матрицы – ДНК или РНК – различают ДНК-зависимую и РНК-зависимую РНК-полимеразу. РЕГУЛЯТОРНЫЕ БЕЛКИ - группа белков, участвующих в регуляции разл. биохим. процессов. Подавляющее большинство таких Р. б. функционирует на уровне транскрипции (синтез матричных РНК, или мРНК, на ДНК-матрице) и отвечает за активацию или подавление синтеза мРНК.
75Один из мощных ингибиторов белкового синтеза – пуромицин. Он представляет собой аналог аминоацил-тРНК адениловой кислоты и поэтому легко взаимодействует с центром пептидил-тРНК с образованием пептидил-пуромицина. Пептидил-пуромицин тормозит элонгацию пептидной цепи, вызывая обрыв реакции, т. е. преждевременную терминацию синтеза белка. Он тормозит синтез всех типов клеточной РНК, особенно мРНК. Данное свойство объясняется тормозящим влиянием актиномицина D на ДНК-зависимую РНК-полимеразу, поскольку он связывается с остатками дезоксигуанозина цепи ДНК, выключая матричную функцию последней. Другим антибиотиком, также тормозящим синтез клеточной РНК, является используемый при лечении туберкулеза рифамицин. Этот препарат тормозит ДНК-зависимую РНК-полимеразу, связываясь с ферментом. Наиболее чувствительной к нему оказалась бактериальная РНК-полиме-раза.
76.???
77В мочевину входит азот, находящийся в белке. Синтез мочевмны на 90 % происходит в печени. Должна переноситься с кровью в почки. Нарушение конечного этапа метаболизма азотосодержащих веществ связана с печенью и с почками. При этом поражении печени приводит к задержки мочевины. В крови повышается содержание низкомолекулярных азотосодержащих веществ (продукциональная гиперазотоеммия) При поражение печени развивается резестентная гиперазотоеммия и в крови резко повышаеться содержание мочевины.
78ЛИПИДЫ (от греч. lipos - жир)- жироподобные в-ва, входящие в состав всех живых клеток. Иногда к липидам относят любые природные в-ва, извлекаемые из организмов, тканей или клеток. В нек-рых случаях липиды рассматривают как производные жирных к-т. В соответствии с хим. строением различают три осн. группы липидов: 1) жирные к-ты и продукты их ферментативного окисления, 2) глицеролипиды (содержат в молек) липиды, не содержащие в молекуле остаток глицерина (за исключением соед., входящих в первую группу). В первую группу входят наряду с жирными к-тами простагландины и др. гидроксикислоты; во вторую - моно-, ди - и триглицериды и большинство фосфолипидов; в третью группу входят сфинголипиды, стерины и воски. По др. классификации липиды подразделяют на нейтральные липиды, фосфолипиды и гликолипиды. Нейтральные липиды (жиры) представляют собой форму депонирования метаболической энергии. Фосфолипиды, гликолипиды и стерины - структурные компоненты биологических мембран; оказывают влияние на множество мембранных процессов, на транспорт ионов, активность мембраносвязанных ферментов, межклеточные взаимод. и рецепцию. Некоторые гликолипиды - рецепторы или корецепторы гормонов, токсинов, вирусов и др. Фосфатидилинозиты участвуют в передаче биол. сигналов. Эйкозаноиды - высокоактивные внутриклеточные регуляторы, межклеточные медиаторы, участвующие в развитии защитных реакций и воспалит. процессов.
79Жиры — сложные эфиры трехатомного спирта глицерина и высших или средних жирных кислот, главная составная часть животных жиров и растительных масел, присутствуют во всех животных и растительных тканях, в питании человека являются одним из основных пищевых веществ. Физико-химические свойства Жиров определяются природой остатков жирных кислот в их молекуле, содержащие значительные количества насыщенных жирных кислот (пальмитиновой, стеариновой и др.), имеют более высокую температуру плавления; Жиры, в состав которых входит много моно - и полиненасыщенных жирных кислот, при обычной температуре находятся в жидком состоянии и называются маслами. Растительные масла, характеризующиеся большим содержанием полиненасыщенных жирных кислот (льняное, маковое, тунговое масло), известны под названием высыхающих масел, т. к. под действием кислорода воздуха они полимеризуются и затвердевают. Биологическая полноценность Жиров. определяется наличием в их составе жирорастворимых витаминов A, D и Е, а также легкостью всасывания в желудочно-кишечном тракте. Жиры. хорошо растворяются в органических растворителях — бензоле, эфире, горячем спирте и не растворяются в воде. Нейтральные жиры вступают во все химические реакции, свойственные сложным эфирам (продуктам замещения атомов водорода в ОН-группах минеральных или карбоновых кислот). Для получения из растительных масел жиров более твердой консистенции, использующихся в качестве жировой основы при производстве маргаринов, применяется гидрогенизация (гидрирование), т. е. насыщение молекул этих масел водородом.
80В первую фазу в-окисления, в течение четырех реакций удаляются ацетил-Кофермент А от карбоксильного конца насыщенных ацил-Кофермента А: 1) дегидрирование б и в-атомов углерода 2) гидрирование с образованием транс - двойных связей 3) дегидрирование образованного L-в-гидроксиацил-КоА 4) КоА-зависимое расщепление с образованием в-кетоацил-КоА и укороченного на 2 атома углерода ацил-КоА. Во вторую фазу окисления жирных кислот, ацетил-КоА окисляется с образованием CO2. Большая часть свободной энергии окисления жирных кислот запасается в виде АТФ.
81. Фосфолипимды — сложные липиды, сложные эфиры многоатомных спиртов и высших жирных кислот. Содержат остаток фосфорной кислоты и соединенную с ней добавочную группу атомов различной химической природы.
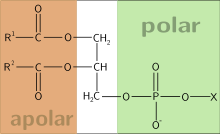 - это фосфолепид.
- это фосфолепид.
Классификация фосфолипидов:
В зависимости от входящего в их состав многоатомного спирта принято делить фосфолипиды на три группы:-глицерофосфолипиды (глицерофосфатиды) - содержат остаток глицерина ;-фосфатидилхолин (лецитин);-фосфатидилэтаноламин (кефалин);-фосфатидилсерин; - кардиолипин;-плазмены или плазмогены(этаноламиновый плазмоген); - фосфосфинголипиды - содержат остаток сфингозина ;-сфингомиелины;
-фосфоинозитиды - содержат остаток инозитола ;-фосфатидилинозитол.
Биологическая роль фосфолипидов-Главный липидный компонент клеточной мембран. Они сопутвуют жирам в пище и служат источником фосфорной кислоты, необходимый для жизни человека
Фосфолипиды — сложные липиды, в которых содержатся жирные кислоты, фосфорная кислота и дополнительная группа атомов, во многих случаях содержащая азот. Они есть во всех живых клетках. Содержатся в нервной ткани, участвуют в транспорте жиров, жирных кислот и холестерина.
Фосфолипиды входят в состав всех клеточных мембран. Между плазмой и эритроцитами происходит обмен фосфолипидами, которые играют важнейшую роль, поддерживая в растворимом состоянии неполярные липиды. Наиболее распространенная группа Фосфолипидов — фосфоглицериды, также к фосфолипидам относятся фосфосфинголипиды и фосфоинозитиды.
Фосфолипиды — амфифильные вещества. Они состоят из полярной «головки», в состав которой входит глицерин или другой многоатомный спирт, отрицательно заряженный остаток фосфорной кислоты и часто несущая положительный заряд группа атомов, и двух неполярных «хвостов» из остатков жирных кислот. Главная особенность фосфолипидов состоит в том, что «головка» у них гидрофильна, а «хвосты» гидрофобны. Это позволяет при нахождении в толще водной среды образовывать бислой — двойной слой фосфолипидных молекул, где гидрофильные головы с обеих сторон соприкасаются с водой, а гидрофобные хвосты упрятаны внутрь бислоя и тем самым защищены от контакта с водой.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 |
