

В процессе гликолиза пищевые вещества подвергаются окислению (дегидрированию) без участия кислорода. Конечным акцептором водорода (окислителем) в этом случае служит молекула какого-либо органического вещества. Энергетическая эффективность гликолиза невелика, и в конечном счете из каждой использованной молекулы глюкозы образуется всего 2 молекулы АТФ.(Аденозинтрифосфат — Универсальный источник энергии для всех биохимических процессов, протекающих в живых системах.)
87. Энергетическая потребность - это полное соответствие пищевой ценности питания энергетическим затратам организма (для поддержания жизни и выполнения работы). Энергетическая ценность выражается в килоджоулях (кДж) или килокалориях (ккал). Вся энергетическая потребность - это совокупность основной, поддерживающей и рабочей потребностей организма человека.
Основной обмен - это количество энергии, необходимое для поддержания жизни организма, который находится в состоянии покоя, то есть утром, лежа в кровати натощак при комфортной температуре. Для мужчин эта величина составляет 1 ккал на 1 кг массы тела в час, а для женщин - 0,9 ккал/кг/час. Все проявления соц активности (работа, трад…)требуют дополнительного количества энергии. В соответствии с характером трудовой деятельности, характером активности население может быть поделено на ряд профессиональных групп:-лица занятые приимущественно умственным трудом(коэффициент физической активности для них равен 1,4);-лица занятые физическим и умственным трудом(коэф. физ. активности равен 1,6) и т. д.
Энергия черпается за счет энергии компонентов пищи, при этом 1г углеводов дает 4,1 ккал,1г белков 4,1 ккал,1г жиров 9,3 кккал.
88. Аденозинтрифосфамт (сокр. АТФ) — нуклеотид, играет исключительно важную роль в обмене энергии и веществ в организмах; в первую очередь соединение известно как универсальный источник энергии для всех биохимических процессов, протекающих в живых системах.
Главная роль АТФ в организме связана с обеспечением энергией многочисленных биохимических реакций. Являясь носителем двух высокоэнергетических связей, АТФ служит непосредственным источником энергии для множества энергозатратных биохимических и физиологических процессов. Все это реакции синтеза сложных веществ в организме: осуществление активного переноса молекул через биологические мембраны, в том числе и для создания трансмембранного электрического потенциала; осуществления мышечного сокращения.
Фосфорилирование — процесс присоединения к какому-либо субстрату остатков фосфорной кислоты. В организме АТФ синтезируется из АДФ, используя энергию окисляющихся веществ
АДФ + H3 PO4(ортофосфорная кислота) + энергия → АТФ + H2 O.
Фосфорилирование АДФ(аденозиндифосфат)возможно двумя способами: субстратное фосфорилирование и окислительное фосфорилирование. Основная масса АТФ образуется на мембранах митохондрий в ходе окислительного фосфорилирования H-зависимой АТФ-синтазой. Субстратное фосфорилирование АТФ не требует участия мембранных ферментов, оно происходит в процессе гликолиза или путём переноса фосфатной группы с других макроэргических соединений.
Реакции фосфорилирования АДФ и последующего использования АТФ в качестве источника энергии образуют циклический процесс, составляющий суть энергетического обмена.
В организме АТФ является одним из самых часто обновляемых веществ, так у человека продолжительность жизни одной молекулы АТФ менее 1 мин. В течение суток одна молекула АТФ проходит в среднем 2000—3000 циклов ресинтеза (человеческий организм синтезирует около 40 кг АТФ в день), то есть запаса АТФ в организме практически не создаётся, и для нормальной жизнедеятельности необходимо постоянно синтезировать новые молекулы АТФ.
Катаболимзм - процесс метаболического распада, разложения на более простые вещества или окисления какого-либо вещества, обычно протекающий с высвобождением энергии в виде тепла и в виде АТФ.
89. Цимкл трикарбомновых кисломт— центральная часть общего пути катаболизма, циклический биохимический аэробный процесс, в ходе которого происходит превращение двух - и трёхуглеродных соединений, образующихся как промежуточные продукты в живых организмах при распаде углеводов, жиров и белков, до CO2. При этом освобождённый водород направляется в цепь тканевого дыхания, где в дальнейшем окисляется до воды, принимая непосредственное участие в синтезе универсального источника энергии — АТФ. Это ключевой этап дыхания всех клеток, использующих кислород, центр пересечения множества метаболических путей в организме. Кроме значительной энергетической роли циклу отводится также и существенная пластическая функция, то есть это важный источник молекул-предшественников, из которых в ходе других биохимических превращений синтезируются такие важные для жизнедеятельности клетки соединения как аминокислоты, углеводы, жирные кислоты и др. Образовавшийся в результате окислительного декарбоксилирования пирувата в митохондриях ацетил-КоА вступает в цикл Кребса. Данный цикл происходит в матриксе митохондрий и состоит из восьми последовательных реакций (рис. 10.9). Начинается цикл с присоединения ацетил-КоА к оксалоацетату и образования лимонной кислоты (цитрата). Затем лимонная кислота (шестиуглеродное соединение) путем ряда дегидрирований (отнятие водорода) и двух декарбоксилирований (отщепление СО2) теряет два углеродных атома и снова в цикле Кребса превращается в оксалоацетат (четырехуглеродное соединение), т. е. в результате полного оборота цикла одна молекула ацетил-КоА сгорает до СО2 и Н2О, а молекула окса-лоацетата регенерируется.
90. Дыхательная цепь является частью процесса окислительного фосфорилирования (см. с. 126). Компоненты дыхательной цепи катализируют перенос электронов от НАДН + Н+ или восстановленного убихинона (QH2) на молекулярный кислород. Из-за большой разности окислительно-восстановительных потенциалов донора (НАДН + Н+ и, соответственно, QH2) и акцептора (О2) реакция является высокоэкзергонической (см. с. 24). Большая часть выделяющейся при этом энергии используется для создания градиента протонов (см. с. 128) и, наконец, дляобразования АТФ с помощью АТФ-синтазы. 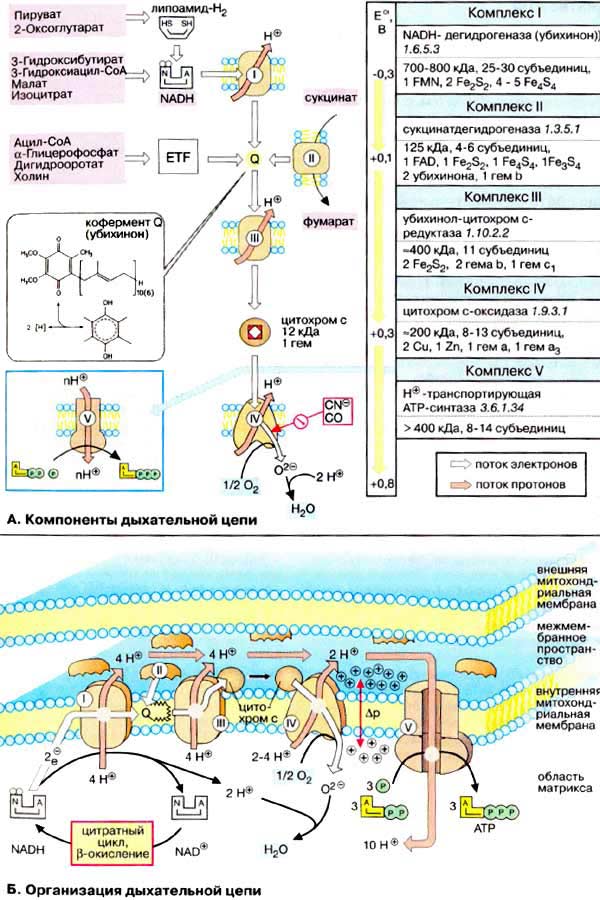
Дыхательная цепь включает три белковых комплекса (комплексы I, III и IV), встроенных во внутреннюю митохондриальную мембрану, и две подвижные молекулы-переносчики — убихинон (кофермент Q) и цитохром с. Сукцинатдегидрогеназа, принадлежащая собственно к цитратному циклу, также может рассматриваться как комплекс II дыхательной цепи. АТФ-синтаза (см. с. 144) иногда называется комплексом V, хотя она не принимает участия в переносе электронов.
Комплексы дыхательной цепи построены из множества полипептидов и содержат ряд различных окислительно-восстановительных коферментов, связанных с белками (см. сс. 108, 144). К ним принадлежат флавин [ФМН (FMN) или ФАД (FAD), в комплексах I и II], железо-серные центры (в I, II и III) и группы гема (в II, III и IV). Детальная структура большинства комплексов еще не установлена.
Электроны поступают в дыхательную цепь различными путями. При окислении НАДН + Н+ комплекс I переносит электроны через ФМН и Fe/S-центры на убихинон. Образующиеся при окислении сукцината, ацил-КоА и других субстратов электроны переносятся на убихинон комплексом II или другой митохондриальной дегидрогеназой через связанный с ферментом ФАДН2 или флавопротеин (см. с. 166), При этом окисленная форма кофермента Q восстанавливается в ароматический убигидрохинон. Последний переносит электроны в комплекс III, который поставляет их через два гема b, один Fe/S-центр и гем с1 на небольшой гемсодержащий белок цитохром с. Последний переносит электроны к комплексу IV, цитохром с-оксидазе. Цитохром с-оксидаза содержит для осуществления окислительно-восстановительных реакций два медьсодержащих центра (CuA и CuB) и гемы а и а3, через которые электроны, наконец, поступают к кислороду. При восстановлении О2 образуется сильный основной анион О2-, который связывает два протона и переходит а воду. Поток электронов сопряжен с образованным комплексами I, III и IV протонным градиентом. Перенос протонов комплексами I, III и IV протекает векторно из матрикса в межмембранное пространство. При переносе электронов в дыхательной цепи повышается концентрация ионов H+, т. е. понижается значение рН. В интактных митохондриях по существу только АТФ-синтаза (см. с. 144) позволяет осуществить обратное движение протонов в матрикс. На этом основано важное в регуляторном отношении сопряжение электронного переноса с образованием АТФ (см. с. 146).
Как уже упоминалось, все комплексы с I по V интегрированы во внутренней мембране митохондрий, тем не менее обычно они не контактируют друг с другом, так как электроны переносятся убихиноном и цитохромом с. Убихинон благодаря неполярной боковой цепи свободно перемещается в мембране. Водорастворимый цитохром с находится на внешней стороне внутренней мембраны.
Основное количество молекул АТФ вырабатывается по способу окислительного фосфорилирования на последней стадии клеточного дыхания: в электронтранспортной цепи. Здесь происходит окисление НАД∙Н и ФАДН2, восстановленных в процессах гликолиза, в-окисления, цикла Кребса и т. д.. Энергия, выделяющаяся в ходе этих реакций, благодаря цепи переносчиков электронов, локализованной во внутренней мембране митохондрий (у прокариот — в цитоплазматической мембране), трансформируется в трансмембранный протонный потенциал. Фермент АТФ-синтаза использует этот градиент для синтеза АТФ, преобразуя его энергию в энергию химических связей. Подсчитано, что молекула НАД∙Н может дать в ходе этого процесса 2.5 молекулы АТФ, ФАДН2 — 1.5 молекулы.
Конечным акцептором электрона в дыхательной цепи аэробов является кислород.
Разобщение осфорилирования-состояние в митохондриях, когда процесс окисления и выделения свободной энергии протекает, а синтез АТФ не происходит. Возможно при воздействии некоторых веществ, нарушении структуры внутренних мембран митохондрий (например, при интенсивной физической разминке) и других факторах
91. Микросомальное окисление - совокупность реакций первой фазы биотрансформации ксенобиотиков и эндогенных соединений, катализирующихся ферментными системами мембран эндоплазматического ретикулума гепатоцитов при участии цитохрома Р-450. При дифференциальном центрифугировании эндоплазматический ретикулум оказывается в микросомальной фракции, поэтому эти реакции получили название микросомальных, а соответствующие ферменты - микросомальных оксигеназ.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 |
