
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Чтобы проанализировать далее признаки генной модификации на геномном уровне, мы непосредственно упорядочивали продукты ПЦР-амплификации интрона 22/экзон 23 соединения от клеток контроля и MDX3-инфицированного-вирусом. Преобразование G-T могло быть обнаружено после одного раундаПЦР - амплификации, и подтверждено после второго раунда амплификации, только в MDX3-инфицированных-вирусом клетках (Рис. 2C). Эти результаты были воспроизведены в 10 независимых экспериментах и были подтверждены анализом последовательности, используя и простые и обратные инициаторы. Эти результаты подтверждают изменение основания нацеленным химеропластом в субпопуляции инфицированных вирусом клеток на геномном уровне.
Экспрессия белка дистрофина в химеропласт-леченных mdx мышечных клетках
Восстановление экспрессии дистрофина было оценено спустя 3 недели после трансфекции и 24–96 часов после индукции дифференцировки как ранее описано (20). Вестерн-блоттинг используя антитела к области белка дистрофина продемонстрировал по крайней мере два различных белковых продукта, молекулярные массы которых были меньше чем дистрофин во всю длину (Рис. 3A). Антитела, направленные к C-терминальной области, подтвердили экспрессию этих двух продуктов, но также и показало наличие третьего белкового продукта еще более низкой молекулярной массы (Рис. 3A). Основанный на специфичности антител, большие продукты содержали экзон 31/32 область белка, тогда как меньший продукт не содержал. Возможно получить только грубое приближение молекулярных масс этих продуктов. Все белки двигались быстрее чем дистрофин или утрофин, таким образом устанавливая 400 kDa как верхний предел их размеров, и все белки двигались медленнее чем самый высокий маркер молекулярной массы 200 kDa, таким образом устанавливая более низкий предел. Два более тяжелых белка (Рис. 3A) молекулярной массы были ближе к 400 kDa концам спектра, и более низкой молекулярной массы (признанный только антителом против C-терминальной области белка) двигалась ближе к 200 концам kDa. Эта структура белков дистрофина была получена в пяти независимых экспериментах и была подтверждена иммунопреципитацией. Кроме того эта структура экспрессии присутствовала в культурах мышечных клеток, полученных из MDX3-инфицированных-вирусом миобластов, которые поддерживались в течение 3 месяцев после трансфекции, подтверждая, что эффект лечения химеропластом был устойчиво унаследован.
Большая версия изображения
In this page
In a new window
Download as PowerPoint Slide
Рисунок 3.

Мы ранее использовали вестерн-блот-анализ, чтобы количественно определить уровень экспрессии дистрофина в культурах mdx клеток, инфицированных вирусом с химеропластом, таким образом обеспечивая меру эффективности конверсии гена (20). В этом текущем исследовании, однако, наличие многократных связок на вестерн-блоттингах сделало эту меру из эффективности более трудной. Как альтернативный подход, мы построили стандартную кривую, используя смесь экстрактов из C57 и mdx мышечных волокон, чтобы определить предел нашей способности обнаружить белковую экспрессию дистрофина вестерн-блот-анализом. Согласно этим исследованиям, мог быть обнаружен дистрофин, пока процент клеток с экспрессией дистрофина был больше чем 1 %. Таким образом мы пришли к заключению, что эффективность замены одного основания установленая MDX3 химеропластом, была в диапазоне 1-5 %. Этот диапазон подобен тому, что ранее сообщалось для химеропласта, разработанного, чтобы исправить mdx мутацию (20,21) ), и находится в диапазоне химеропласт-опосредованного преобразования, показанного другими (19,23,26).
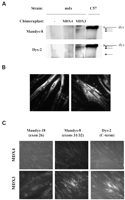
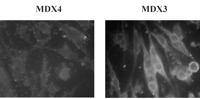
Экспрессия некоторых форм белка дистрофина в субпопуляции MDX3-инфицированных-вирусом клеток была также подтверждена иммунным окрашиванием. Последовательно с результатами вестерн-блоттинга, мы наблюдали группы дистрофин-положительных волокон после дифференцирования только в MDX3-инфицированных-вирусом клетках (Рис. 3B). Иммунное окрашивание леченных клеток с определенными для экзона анти-дистрофиновыми антителами подтвердило наличие внутренне измененных форм дистрофина в мышечных волокнах, полученных из MDX3-инфицированных-вирусом клеток (Рис. 3C), тогда как культуры мышечной волокон, полученные из неинфицированных вирусом клеток или из MDX4-инфицированных-вирусом клеток, не окрашивали положительно ни одним из антител. В культурах мышечных волокон, полученных из MDX3-инфицированных-вирусом клеток, групп мышечных волокон, положительно окрашенных антителами против C-терминальной области и экзона 31/32 области дистрофина, но ни одно из мышечных волокон, окрашенных положительно антителом против экзона 26 (Рис. 3C). Эти данные согласуются с данными о вестерн-блоттинге, указывая, что изменение интрона 22/экзон 23 места соединения заканчивается многократным пропуском экзонов, не только экзона 23 (Рис. 3A). Многократный пропуск экзона, как наблюдали другие исследователи, использующие антисмысловые олигонуклеатиды, вмешался в интрон 22/экзон 23 соединения (9).
Восстановление экспрессии б-дистрогликана
Как было показано экспрессия б-дистрогликана (б-DG), большого компонента дистрофин-гликопротеинового комплекса снижено до 80-90% у mdx мышей из-за нехватки дистрофина (27). Мы использовали иммунное окрашивание, чтобы определить, могли ли белки дистрофина, произведенные разрушением интрона 22/экзон 23 соединения, восстановить экспрессию б-DG. Иммунное окрашивание MDX3-инфицированных-вирусом культур, которые дифференцировались в течение 4 дней, подтвердило локализацию б-DG в мембране сарколеммы в группах мышечных волокон (Рис. 4). Эти результаты ясно демонстрируют, что по крайней мере один из произведенных белков был функционален, совместим с функциональными свойствами спроектированных дистрофинов (28).
Большая версия изображения:
In this page
In a new window
Download as PowerPoint Slide
Рисунок 4.
Обнаружение измененных продуктов сплайсинга, вызванных MDX3

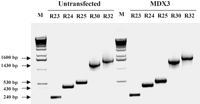
Основанное в иммунном окрашивании и иммуноблот-анализе использование определенных для экзона антител, мы идентифицировали продукты измененного сплайсинга из-за пропуска экзона 23. Мы использовали анализ RT–PCR, чтобы идентифицировать продукты измененного сплайсинга, вызванные разрушением интрона 22/экзон 23 места соединения в клетках, которые были поддержаны в культуре спустя 2 недели после трансфекция химеропласта. Мы проектировали определенные инициаторы,


перекрывающие экзон 22 до экзона 32 из гена дистрофина. Продукты, представляющие транскрипты во всю длину, были амплифицированны от невылеченных и химеропласт-леченных клеток, используя эти инициаторы (Рис. 5). Мы были неспособны обнаружить любые альтернативно соединенные транскрипты в этой области в клетках, инфицированных вирусом с целенаправленным химеропластом (Рис. 5). Отсутствие альтернативных продуктов соединения было также подтверждено введением дополнительного шага амплификации RT–PCR, используя внутренние инициаторы (данные, не показанны).
Большая версия изображения:
In this page
In a new window
Download as PowerPoint Slide
Рисунок 5.
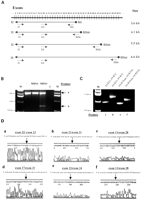
Мы поэтому расширяли наше исследование, используя инициаторы, которые охватили участок от экзона 9 до экзона 50 (Рис. 6A), таким образом покрывая почти 50 % всей ДНК дистрофина. Вторая амплификация ПЦР была выполнена, используя пары инициатора (см. Материалы и Методы и Рис. 6A), расположенный в области транскрипта ддистрофина, усиленного при первом шаге. Мы наблюдали пять продуктов (маркировал ‘b’, ‘c’, ‘d’, ‘e’ и ‘f’) в MDX3-инфицированных-вирусом клетках (Рис. 6B и C), и эта структура также наблюдалась в клетках, которые поддерживались в культуре в течение 3 месяцев после трансфекции. Ни один из этих продуктов никогда не обнаруживался в неинфицированных вирусом клетках или клетках, инфицированных вирусом с контрольным химеропластом. Прямое отщепление этих продуктов показало, что они представляли пять различных разновидностей сайтов сплайсинга, каждый пропускающий экзон 23 плюс дополнительные экзоны и вверх по течению и вниз по течению интрона 22/экзон 23 соединения, с 26 экзонами (экзоны 12–37) пропускаемый в самом маленьком продукте (Рис. 6C и D). Три из пяти разновидностей сайтов сплайсинга (продукты ‘c’, ‘d’ и ‘f’) были в структуре (Таблица 1). Предсказанные последовательности аминокислот из этих транскриптов в структуре включили бы белки между 271 и 350 kDa (Таблица 1), совместимые с диапазоном, предложенным вестерн-блот-анализом (Рис. 3A).
Большая версия изображения:
In this page
In a new window
Download as PowerPoint Slide
Рисунок 6.
Primers | Full-length product size predicted | Alternate product size observed | Product label (Fig. 6) | Altered splicing | Reading frame | Predicted protein size |
F19–R32 | 2100 bp | 600 bp | b | 21/31 | Out-of-frame | - |
F9–R32 | 3410 bp | 900 bp | с | 11/28 | In-frame | 331 kDa |
F9–R32 | 3410 bp | 1300 bp | d | 17/31 | In-frame | 346 kDa |
F9–R35in | 3930 bp | 700 bp | e | 12/34 | Out-of-frame | — |
F9–R43in | 5100 bp | 1100 bp | f | 11/38 | In-frame | 271 kDa |
Таблица 1.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
