
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Для качественного исследования микрофлоры носовой полости и кишечника проводили посевы на ряд элективных и дифференциальных сред. Стрептококки выделяли на среде Гарро и КАМП-методом. Для выделения стафилококков использовали кровяной солевой МПА. Из чистой культуры, выращенной на МПА, ставили реакции на плазмокоагуляцию, фибринолизин, лецитиназу, ДНК-азу и скрытую гемолитическую активность. Из биохимических свойств определяли разжижение желатины, коагуляцию молока, ставили реакции на маннит, лактозу, сахарозу, аммиак, сероводород. Клостридии культивировали на мясо-пептонно-печеночном бульоне (МППБ) Китта-Тароцци, плотной среде Вильсона Блера, глюкозо-кровяном агаре Цейслера. Выделение бифидобактерий проводили посевом больших разведений фекалий в среду Блаурокка. Лактобациллы определяли на среде МРС. Для выделения бактерий семейства кишечных материал засевали на среды Эндо, Левина, МПА, МПБ. Чистую культуру эшерихий типировали в реакции агглютинации. Для выделения протея материал засевали в конденсационную воду свежескошенного агара. Выделение микроскопических грибов осуществляли на средах Сабуро и Чапека. Результаты переводили (при посеве из кишечника) в десятичные логарифмы и устанавливали относительное соотношение различных групп микроорганизмов.
Содержание трийодтиронина (Т3), свободного тироксина (сТ4) и тиреотропного гормона (ТТГ) определяли иммуноферментным методом с использованием стандартных тест-наборов в соответствии с прилагаемыми инструкциями фирмы «Алкор-Био». С этой целью применяли соответствующие наборы реагентов: для Т3 –«Тироид ИФА-трийодтиронин-01», сТ4 - «Тироид ИФА-свободный Т4», ТТГ - «Тироид ИФА-ТТГ-1».Содержание йода определяли на атомно-абсорбционном спектрометре «Квант-ZЭТА» по стандартной методике.
Полученный цифровой материал подвергнут статистической обработке методами вариационной статистики с проверкой достоверности результатов с помощью критерия Стьюдента с использованием соответствующих стандартных компьютерных программ (,1980).
2.2. Результаты собственных исследований
2.2.1. Естественная резистентность, фагоцитоз свиней при йодной недостаточности и разных методах ее коррекции
В сыворотке крови животных 2-8-й опытных групп, с йодной недостаточностью в организме, бактерицидная активность к началу опытов была снижена.
Показатель бактерицидной активности сыворотки крови свиней второй группы в процессе опыта, имел тенденцию к дальнейшему снижению. На 10, 30, 60 и 120-ые дни он понизился, по сравнению с контрольным уровнем, в 1,7; 2,08; 2,41и 2,63 раза. Введение йодида калия per os и аэрозольно (третья и четвертая группы) способствовало умеренной активизации ее. Более выраженным этот процесс был в организме животных четвертой группы. Значительная активизация бактерицидной активности сыворотки крови отмечалась у животных пятой и шестой групп. Максимального уровня бактерицидная активность сыворотки крови достигла у поросят седьмой и, особенно, восьмой групп. Она превысила фоновый уровень к 10, 30, 60 и 120-ым дням опыта по седьмой группе соответственно в 1,17;1,42; 1,54 и 1,47 раза, по восьмой – в 1,3; 1,65; 1,82 и 1,73 раза. До конца опыта описываемый показатель в сыворотке крови поросят восьмой группы был выше их значения в контрольной и других опытных группах.
Результаты исследований фагоцитарной активности лейкоцитов крови представлены на рисунке 2.
Данные по изучению динамики лизоцимной активности сыворотки поросят приведены в таблице 2.
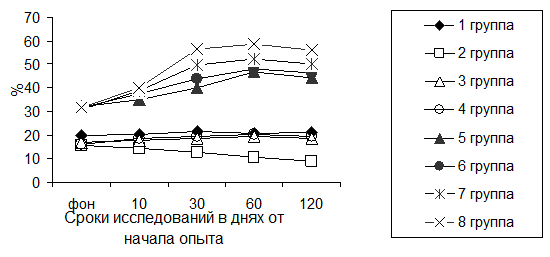
Рисунок 2 Динамика фагоцитарной активности лейкоцитов крови поросят при йодной недостаточности и разных методах ее коррекции
2.2.2 Т – и В - системы иммунитета свиней при йодной недостаточности и разных методах ее коррекции
Содержание Т-Е-РОК - лимфоцитов в крови поросят с выраженной йодной недостаточностью было снижено на 8,9 – 11,0% и на 0,4 – 0,7*109 /л. Т-Е-РОК-лимфоциты в крови животных 2-ой группы имели тенденцию к дальнейшему снижению и к 120- му дню опыта эта разница была на 22,6%. Т-Е-РОК - лимфоциты в крови поросят 3-8-й опытных групп, в процессе эксперимента, постепенно повышались, по сравнению с фоновым показателем. Умеренным этот процесс был в крови животных 3 и 4-ой групп. Количество Т-Е-РОК-лимфоцитов в крови поросят 5 и 6-ой групп имело более выраженную степень повышения. Самого высокого показателя Т-Е-РОК - лимфоциты достигли в крови поросят 7 и, особенно, 8-ой групп. Здесь их содержание было выше, по сравнению с фоновым уровнем, к 10, 30, 60 и 120-ым дням опыта на 5,9; 8,8; 12,8; 10,5% и на 8,4; 12,4; 16,9 и 12,5%.
Таблица 2 Динамика изменения лизоцимной активности сыворотки
крови поросят (M±m, в %)
Группа животных | Фон | Сроки исследований в днях от начала опытов | |||
10 | 30 | 60 | 120 | ||
1 | 19,80±0,73 | 20,20±0,80 | 21,70±0,73 | 20,60±1,08 | 20,90±0,87 |
2 | 15,40±0,51* | 14,30±0,66** | 12,80±0,66*** | 10,60±0,40*** | 8,70±0,30*** |
3 | 16,80±0,58* | 17,80±0,86* | 18,40±0,51* | 19,30±0,62 | 18,70±0,62 |
4 | 15,90±1,03* | 18,60±0,51 | 19,60±0,75 | 20,10±0,71 | 19,20±1,02 |
5 | 16,80±0,66* | 19,40±0,51 | 20,60±0,68 | 21,60±0,93 | 20,00±0,71 |
6 | 16,20±0,37* | 19,90±0,46 | 21,00±0,95 | 22,00±0,71 | 20,50±0,92 |
7 | 16,40±1,08* | 20,60±0,93 | 22,30±1,16 | 25,60±1,03** | 24,80±0,97** |
8 | 16,00±1,05* | 21,20±0,73 | 24,50±0,89** | 27,00±0,71*** | 25,20±1,02** |
Примечание: при Р<0,05 - *, Р<0,01 - *, Р<0,001 - *** - разница достоверна.
Йодная недостаточность способствовала снижению продукции В-ЕАС - клеток в организме свиней. Фоновый уровень В - лимфоцитов в крови поросят 2-8-ой групп был ниже, чем в контроле, на 6,8 – 7,8%.У животных 2-ой группы регистрировалось дальнейшее снижение уровня В - клеток в крови: к 10, 30, 60 и 120-ым дням опыта на 7,2; 8,2; 7,9 и 8,7%.Содержание В-ЕАС-лимфоцитов в крови свиней опытных групп повышалось по срокам опыта. Максимальная реакция В - лимфоцитов отмечалась в крови свиней 7 и 8-ой групп.
Содержание Т-Е-РОК-клеток в лимфоузле поросят 2-8-й опытных групп, к началу опытов (фон), было снижено. В лимфатическом узле поросят 2-й группы процесс затормаживания активности Т-Е-РОК-лимфоцитов прогрессировал по периодам опыта. К 10, 30, 60 и 120-ым дням эксперимента их уровень был ниже контрольного показателя на 3,5; 10,6; 12,0; и 14,4%. Т-Е-РОК-лимфоциты в лимфоузле животных 3 и 4-й групп до 30-го дня умеренно повышались в количестве и достигли к этому сроку контрольного уровня. В лимфатическом узле свиней 5 и 6-й групп количество Т-Е-РОК - лимфоцитов достигло контрольного значения к 10-му дню опыта. Максимальный уровень Т-Е-РОК-лимфоцитов регистрировался в лимфоузле животных 8-й группы. Они превысили фон и контроль к 10,30 60 и 120-ым дням на 6,3 и 4,1%; 13,4 и 9,4%; 15,0 и 11,4%; 12,7 и
9,3%. Незначительно уступали им показатели животных 7-ой группы.
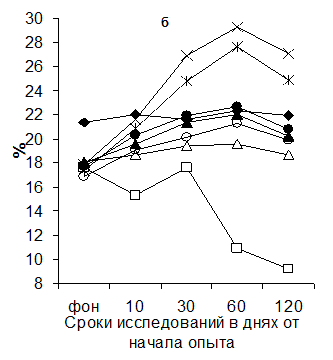 Данные по исследованию динамики Т - супрессоров и Т-хелперов в крови поросят приведены на рисунке 3.
Данные по исследованию динамики Т - супрессоров и Т-хелперов в крови поросят приведены на рисунке 3.
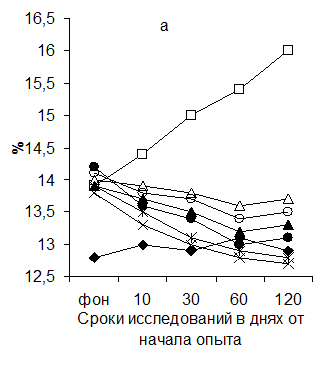
Рисунок 2 Динамика содержания Т-супрессоров (а) и Т-хелперов (б) в крови поросят при йодной недостаточности и разных методах коррекции
Рис.3 Динамика Т-супрессоров и Т-хелперов в крови поросят
(обозначения те же, что и на рис.1)
Данные по изучению динамики В-ЕАС-лимфоцитов в лимфоузле и селезенке животных приведены на рисунке 4.
Фоновый уровень иммунокомпетентных Т - клеток в селезенке свиней, с нарушенным йодным обменом был снижен на 6,7 – 8,0 млн. и 13,6 – 17,6 млн.
В органе животных 2-й группы процесс снижения активности Т - лимфоцитов, по сравнению с фоновым и контрольным показателями, прогрессировал по срокам опыта. Умеренная активизация Т-лимфоцитов наблюдалась в селезенке свиней 3 и 4-й групп. Более выраженным этот процесс был в селезенке животных 5 и 6-й групп. Самый интенсивный процесс активизации Т-Е-РОК-лимфоцитов отмечался в селезенке свиней 7 и 8-й групп. Их уровень повысился, по сравнению с контролем, к 10, 30, 60 и 120-ым дням опыта на 4,0 и 6,8%; 9,9 и 14,1%; 10,1 и 12,5%; 8,8 и 11,2%.
Фоновый уровень Т-Е-РОК - лимфоцитов в тимусе поросят с йодной недостаточностью был снижен на 159,7-170,2 млн. Изменения в тимусе животных 3-8-й опытных групп были направлены на повышение их уровня. Этот процесс имел не одинаковую степень выраженности по группам. Показатели животных 3 и 4-й групп не достигали контрольного уровня. Интенсивнее увеличение Т-Е-РОК - клеток наблюдалось в тимусе поросят 5 и 6-й групп. Однако здесь они также не достигли контрольных цифр. Содержание Т-Е-РОК - лимфоцитов в тимусе поросят 7 и 8-й групп с 30-го дня опыта было выше, чем в контроле.
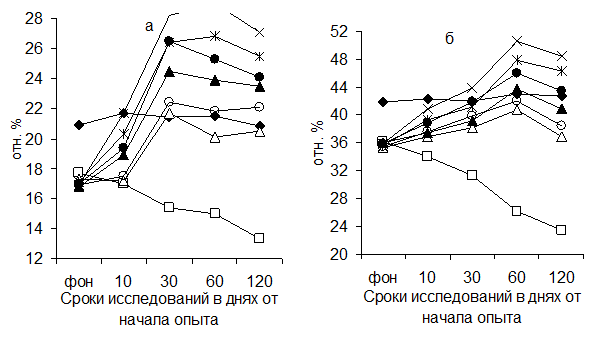
Рисунок 4 Динамика В-ЕАС-лимфоцитов в глубоком паховом лимфатическом узле (а) и селезенке (б) поросят (обозначения те же, что и на рис.1)
2.2.3 Мононуклеарные клетки периферической крови,
иммунофенотипированные моноклональными антителами,
иммуноглобулины сыворотки крови поросят при йодной
недостаточности и разных методах ее коррекции
В крови животных с нарушенным йодным балансом в организме наблюдалось снижение МКА серии ИКО - 124. Их значение в крови поросят 2-й группы, в процессе опыта, имело тенденцию к дальнейшему снижению. У животных 3-8-й опытных групп происходили изменения в сторону их повышения. Максимальное содержание лимфоцитов, реагирующих с МКА серии ИКО - 124 выделялось из крови поросят 7-й и, особенно, 8-й групп. Здесь их уровень увеличился, по сравнению с исходным фоновым показателем, на 10, 30, 60 и 120-ые дни опыта на 4,9 и 6,0%; 7,3 и 8,9%; 8,2 и 11,1%; 7,7 и 10,3%. Подобным образом изменялась динамика МКА серии ИКО- 10.
Результаты исследования изменения содержания в сыворотке крови свиней иммуноглобулинов класса G представлены в таблице 3.
Фоновый уровень Ig А в сыворотке крови животных с йодной недостаточностью был снижен на 0,9 – 1,03 мг/мл. У свиней 2-й группы они уступали контролю к 10, 30, 60 и 120-ым дням на 1,08; 0,96; 1,08 мг/мл и 1,46 мг/мл. В сыворотке крови поросят 3-8-й групп, в процессе опытов, наблюдалось динамичное повышение Ig А. Самый высокий уровень Ig А регистрирован в сыворотке крови свиней 7 и, особенно, 8-й групп.
Таблица 3 Динамика изменения содержания Ig G в сыворотке крови поросят
(M±m, в мг/мл)
Группа животных | Фон | Сроки исследований в днях от начала опытов | |||
10 | 30 | 60 | 120 | ||
1 | 20,80±0,58 | 21,20±0,86 | 21,90±0,87 | 22,00±1,18 | 21,80±0,80 |
2 | 18,60±0,75* | 17,60±1,17* | 17,20±0,73** | 15,30±0,77*** | 15,40±0,68*** |
3 | 17,90±0,56* | 18,80±0,37* | 19,20±0,46 | 20,30±0,77 | 20,00±0,32 |
4 | 18,40±0,99* | 18,90±0,71* | 19,90±0,64 | 21,20±1,02 | 20,20±1,16 |
5 | 18,50±0,50* | 19,20±0,41* | 20,80±0,86 | 21,70±0,86 | 21,20±0,58 |
6 | 17,60±0,93* | 19,50±0,74* | 21,40±0,60 | 22,00±0,61 | 21,60±0,97 |
7 | 18,00±0,71* | 19,80±0,66* | 22,90±0,90 | 23,90±0,68* | 23,30±0,37** |
8 | 17,80±0,73* | 20,10±0,51* | 23,40±0,68* | 25,00±1,10*** | 24,60±0,53*** |
Примечание: при Р<0,05 - *, Р<0,01 - **, Р<0,001 - *** - разница достоверна.
2.2.4 Иммуноморфологические перестройки в структурах лимфоидных органов свиней при йодной недостаточности и разных методах ее коррекции
В лимфатическом узле животных контрольной группы на долю коркового плато приходилось 18,4 -20,6%, лимфатических узелков без светлых центров 7,2 – 7,9%, со светлыми центрами 2,9 – 3,7%, мякотных тяжей 17,9-18,9%, паракортикальной зоны 14,9-16,0%. Площадь коркового плато у поросят 2-8-й групп, к началу опытов, была расширена на 4,9-5,9%. В лимфатическом узле животных 2-й группы в процессе опыта наблюдалось дальнейшее ее расширение. У поросят 3-8-й групп в ходе экспериментов отмечалось уменьшение площади коркового плато. Максимальным тот процесс был в лимфоузле поросят седьмой и восьмой групп. Она уступала фону к концу опыта на 6,0 и 6,3%.
При йодной недостаточности отмечалось уменьшение площади лимфатических узелков без светлых центров. В лимфоузле животных 2-й группы данный процесс активно прогрессировал. В опытных группах отмечалось расширение площади лимфоузелков без светлых центров. Максимальной описываемая площадь была у животных 7 и 8-й групп. Данные по исследованию динамики изменения площади, занимаемой лимфатическими узелками со светлыми центрами и паракортикальной зоной глубокого пахового лимфатического узла представлены на рисунке 5.
В лимфатическом узле животных 2-й группы регистрировалось уменьшение площади мякотных тяжей. К 10, 30, 60 и 120-ым дням этот показатель уступал контрольному уровню на 2,9; 5,7; 7,4 и 10,2%. Во всех опытных группах регистрировалось расширение площади мякотных тяжей органа. Максимального показателя они достигли у поросят 7 и 8-й групп.
Результаты морфометрических измерений структурных компонентов тимуса показали, что фоновый показатель площади коркового вещества у поросят 2-8-й групп в начале опытов был меньше, чем в контроле, на 22,2-24,2%. Площадь коркового вещества тимуса поросят 2-й группы подвергалась в процессе опытов дальнейшему уменьшению: она уступала контрольному показателю на 10, 30,
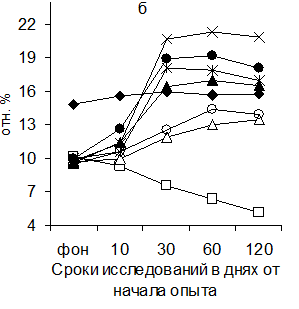
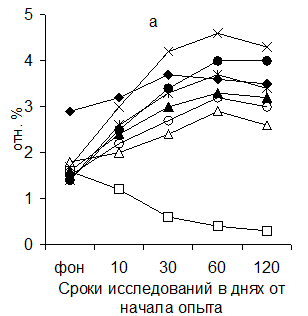
Рисунок 5 Динамика изменения соотношения площади лимфатических узелков со светлыми центрами (а) и паракортикальной Т-зоны (б) в глубоком паховом лимфатическом узле поросят (обозначения те же, что и на рис.1)
60 и 120-ые дни исследований на 24,2; 28,9; 34,2 и 37,3%. Описываемый показатель в тимусе поросят 3-8-й групп, в процессе опытов имел тенденцию к увеличению. Более значительное расширение площади коркового вещества регистрировалось у поросят пятой, шестой, и особенно седьмой и восьмой групп. Максимального значения она достигла у животных этих групп к 60-му дню, превысив фоновое значение на 19,5; 21,0; 23,7 и 26,4%.
Данные морфометрических исследований реакции селезенки представлены в автореферате только по 1, 2 и 8-й группам (таблица 4).
2.2.5 Иммуноцитологические реакции в лимфоидных органах свиней при йодной недостаточности и разных методах ее коррекции
Уровень нейтрофилов в костном мозге свиней 2-й группы к началу опытов был увеличен на 16,09%. При этом показатель миелоцитов превышал контрольную цифру на 2,49%, метамиелоцитов на 5,7%, палочкоядерных нейтрофилов на 5,8%, сегментоядерных на 2,1%. К 60-му дню исследований нейтрофилия в миелограмме свиней 2-й группы прогрессировала. Показатели содержания нейтрофилов в костном мозге свиней 3, 4, 7 и 8-й опытных групп, к 60-му дню опыта, имели тенденцию к снижению в сторону физиологических норм. Однако, они продолжали превышать значение животных 1-й группы, по 3 и 4-й группам на 2,92 и 2,0%, а по 7 и 8-й группам - приблизилось контрольному уровню.
Уровень метамиелоцитов к 60-му дню исследований понизился в 3 и 4-й
Таблица 4 Изменения морфометрических показателей селезенки поросят
Сроки исследований, в днях | Красная пульпа (M±m, в %) | Белая пульпа (лимфатические узелки) (M±m, в %) | ||
Т-зона | В-зона | |||
периваскулярная муфта | без светлых центров | со светлыми центрами | ||
1 группа – контроль, здоровые животные | ||||
10 | 57,40±0,87 | 12,80±0,42 | 14,90±0,49 | 3,60±0,06 |
30 | 58,50±1,04 | 11,90±0,49 | 15,60±0,26 | 3,90±0,25 |
60 | 58,00±2,37 | 13,40±0,87 | 15,00±1,15 | 4,50±0,21 |
120 | 57,90±1,36 | 13,00±1,15 | 14,70±0,65 | 4,30±0,15 |
2 группа – контроль, больные | ||||
10 | 63,20±1,56** | 9,40±0,70** | 10,10±0,59*** | 3,00±0,29 |
30 | 66,70±2,43*** | 8,20±0,57*** | 9,00±0,58*** | 2,20±0,21** |
60 | 72,60±2,35*** | 6,60±0,55*** | 7,30±0,15*** | 1,50±0,15*** |
120 | 79,90±1,73*** | 4,20±0,17*** | 6,20±0,21*** | - |
8 группа - KI аэрозольно + сапропель + пробиотик + прополис | ||||
10 | 63,70±2,33*** | 9,03±0,52*** | 9,80±0,31*** | 2,33±0,15** |
30 | 58,60±0,40 | 11,40±0,31 | 14,60±0,31* | 4,50±0,12 |
60 | 56,00±1,53 | 12,90±1,12 | 15,60±0,87 | 4,80±0,12 |
120 | 53,50±1,26* | 15,70±1,11** | 17,50±1,04** | 5,20±0,23* |
Примечание: при Р<0,05 - *, Р<0,01 - *, Р<0,001 - *** - разница достоверна.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
