

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
При переносе мутантного гена в новую генотипическую среду происходит
значительная модификация корреляционных связей между признаками. У мутантно-сортовых гибридов наблюдалась более тесная связь продуктивности растения с массой 1000 зёрен и количеством колосков и зёрен главного колоса, менее тесная связь была с общей и продуктивной кустистостью. Возможность изменения направления и характера корреляционных связей при мутагенезе представляет не только теоретический, но и практический интерес, так как появляется перспектива изменения нежелательных и создания более благоприятных связей между селекционно-значимыми признаками.
Корреляции количественных признаков у гибридов, полученных с участием мутантов озимых форм. Продуктивность растения в яровых гибридных популяциях, полученных с участием мутантов озимой пшеницы, зависит в большей степени от продуктивной кустистости и массы зерна главного колоса. Вклад дополнительных побегов в общую продуктивность более существенен в благоприятные по увлажнению годы, в условиях же зоны с коротким вегетационным периодом и наличием засухи в первый период роста и развития яровой пшеницы продуктивность обеспечивают в основном главные колосья. В устойчивой положительной корреляции с продуктивностью растения находятся озернённость колоса и масса 1000 зёрен. В благоприятных по увлажнению и температурному режиму условиях среды связь продуктивности растения с массой зерна колоса была средней, в типичных для зоны условиях уровень связи изменялся у одних гибридов и оставался прежним у других, в острозасушливых – уровень связи признаков значительно повышался.
Продуктивность главного колоса в условиях зоны тесно связана с количеством колосков и зёрен в нём, а также с крупностью зерна, причём большее влияние по величине и устойчивое по годам и комбинациям оказала на
массу зерна главного колоса его озернённость. В то же время, достоверной связи между озернённостью колоса и массой 1000 зёрен не установлено, что указывает на возможность сочетания в одном генотипе высокой озернённости и крупности зерна. Отсутствие существенной коррелятивной связи продуктивной кустистости с элементами структуры урожая, меньшая фенотипическая изменчивость продуктивности главного колоса, высокая положительная связь массы зерна колоса и продуктивности растения позволяют предположить, что отбор по продуктивности колоса при оптимальной продуктивной кустистости приведёт к увеличению продуктивности растения.
5.2 Характеристика корреляционных плеяд и их зависимость от мутагенных факторов и условий выращивания
Общий анализ корреляционных плеяд мутантов озимой пшеницы показал наибольшую стабильность структуры плеяд и количества связей в них у Мутанта 85 и Мутанта 33 в разных условиях среды, что свидетельствует о их относительно высокой гомеостатичности. Анализ корреляционных плеяд мутантов озимых форм позволил выделить устойчивую плеяду признаков продуктивности: продуктивность растения - общая кустистость - продуктивная кустистость, коррелирующую по годам на уровне r>0,7, а также слабую, но устойчивую по годам связь продуктивность растения - количество колосков в колосе. Другие связи были более лабильны по годам и объектам исследований. Было выявлено влияние мутагенов на взаимосвязи признаков: большей стабильностью количества и уровня достоверных связей обладают мутанты – 6 корреляций на одном уровне за три года испытаний у мутантов, против 3 у исходного сорта; при общем количестве достоверных связей у Мироновской 808 – 44, Мутанта 85 – 44, Мутанта 88 – 47, что говорит о большей приспособленности мутантов к местным условиям (рис. 9).
Тесная корреляционная связь у яровых сортов во все годы исследований была характерна для признаков общая кустистость - продуктивная кустистость и озернённость главного колоса - масса зерна главного колоса, другие связи подвержены значительным сдвигам, что обусловливает легко заметные перестройки плеяд в разные годы, при этом количество и теснота связей в зависимости от условий вегетации у сортов меняется по-разному.
Корреляционные плеяды озимо-яровых гибридов F1 перестраиваются в изменяющихся условиях сильнее, чем у родительских сортов и гибридов второго поколения. Увеличение корреляционных связей и их лабильность у гетерозиготных гибридов связано с тем, что у гибридов первого поколения проявляются взаимодействия аллелей типа доминирования и сверхдоминирования, фенотипическая реализация которых в сильной степени зависит от влияния внешних условий. Если сравнивать разные поколения гибридов (F1 и F2) по степени лабильности их плеяд, то можно отметить, что у некоторых комбинаций число корреляций в F2 уменьшается, но у большинства гибридов проявляется противоположная тенденция – увеличение количества связей во втором поколении по сравнению с первым.
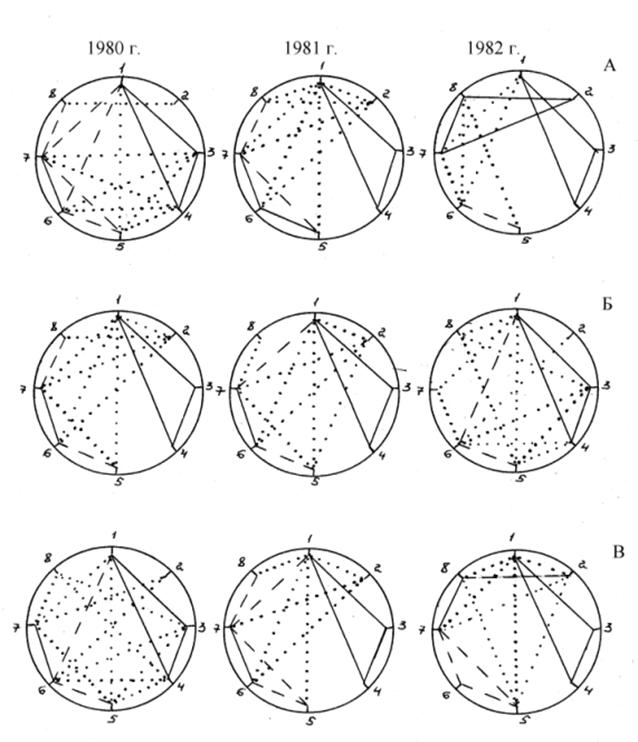
Рис. 9 – Корреляционные плеяды количественных признаков Мироновской 808 (А) и её мутантов Мутанта 85 (Б), Мутанта 88 (В),
где 1 - продуктивность растения, 2 - высота растения, 3 - общая кустистость, 4 - продуктивная кустистость, 5 - количество колосков колоса, 6 - количество зёрен колоса, 7 - масса зерна колоса, 8 - масса 1000 зёрен,
……. – r <0,5; ,5< r <0,7; r >0,7
Во втором поколении гибридных популяций устойчивые тесные корреляционные связи наблюдались между общей и продуктивной кустистостью и озернённостью и массой зерна главного колоса. Корреляционная зависимость продуктивности растения с продуктивностью колоса и его элементами зависела в большей степени от условий выращивания, нежели от генотипов. Устойчивая плеяда признаков озернённость - количество колосков - масса зерна колоса указывает на то, что отбор по количеству колосков в колосе в любых условиях должен привести к повышению озернённости колоса и через неё к повышению массы зерна колоса. Отличаются лабильностью связь массы зерна колоса с массой 1000 зёрен, её уровень может меняться от слабой до тесной в зависимости от условий среды и генотипа.
Большое значение при селекции имеет корреляционная зависимость продуктивности растения и колоса с высотой растения, определяющей устойчивость к полеганию. Корреляционные связи продуктивности растения с высотой у гибридов второго поколения в большинстве случаев слабые, наблюдались единичные случаи среднего уровня связей или их отсутствие. Низкая коррелятивная связь высоты растения с массой зерна колоса наблюдалась во все годы исследований у одних гибридов, средняя – у других, здесь стратегия отбора устойчивых к полеганию высокопродуктивных форм не зависит от условий среды. В третьих комбинациях отборы на устойчивость к полеганию высокопродуктивных форм необходимо проводить с учётом условий выращивания.
В целом, нужно отметить лабильность корреляционных плеяд в зависимости от условий среды. Смена лимитирующих факторов по годам в момент закладки признаков в онтогенезе приводит к переопределению генетических формул количественных признаков, что влечёт за собой сильные сдвиги корреляционных зависимостей.
6 Влияние экологических факторов на донорские способности сортов и мутантов мягкой пшеницы
6.1 Комбинационная способность мутантов озимой пшеницы
Изучение комбинационной способности мутантов озимой пшеницы показало, что мутанты могут служить донорами как отдельных количественных признаков, так и результирующего показателя – урожайности зерна в популяциях яровой и озимой пшеницы. Установлено влияние условий среды на комбинационную способность сортов и мутантов, значения эффектов ОКС непостоянны и изменялись по годам. В большей степени они варьировали по таким признакам, которые сильнее подвержены модифицирующему влиянию среды: продуктивность растений и продуктивная кустистость. По таким признакам, как озернённость, масса зерна главного колоса и масса 1000 зерен эффекты ОКС проявили большую стабильность по годам. Признаки в первой, второй и третьей генерации гибридных популяций в большей степени контролируются действием аддитивных генов мутантов озимой пшеницы, чем яровых родительских форм. Влияние взаимодействия генов родительских форм было значительно меньшее и в большой степени зависело от условий вегетации. Мутанты представляют интерес для селекции яровой и озимой пшеницы по ряду количественных признаков и служат источником генов, увеличивающих общую и продуктивную кустистость (Мутант 85, Мутант 88), количество колосков и зёрен колоса (Мутант 68/8, Мутант 216/16), массу зерна колоса (Мутант 68/8), массу 1000 зёрен (Мутант 68/8, Мутант 88), продуктивность растения (Мутант 85, Мутант 88).
6.2 Комбинационная способность мутантов яровой пшеницы
Изучение комбинационной способности мутантов яровой пшеницы в различных программах скрещиваний показало, что независимо от гибридной популяции и условий среды, в генетическом контроле всех изучаемых признаков преобладали вариансы ОКС, что указывает на значительный вклад в изменчивость признаков генов с аддитивным эффектом действия и позволяет вести отбор нужных форм уже в ранних поколениях гибридных популяций. Достоверность в отдельных комбинациях варианс СКС свидетельствует о роли генов взаимодействия в детерминации признаков, в данном случае отбор следует начинать в более поздних поколениях гибридных популяциий. Проведённые исследования показали, что хемомутанты могут служить донорами количественных признаков: продуктивной кустистости Мутант 562, Мутант 771; количества колосков колоса – Мутант 771; количества зёрен колоса – Мутант 562, Мутант 772, Мутант 777; массы зерна колоса – Мутант 772, Мутант 777, Мутант 562; массы зерна с растения – Мутант 771, Мутант 753; массы 1000 зёрен – Мутант 562, Мутант 771.
6.3 Новые адаптивные формы мягкой пшеницы
Использование мутантного материала позволило создать три сорта озимой пшеницы – Омская озимая (Мироновская 808 ЭИ 0,01%), Сибирская нива (ППГ-186 ЭИ 0,01%) и Омская 3 (Ильичёвка НЭМ 0,05%).
Хемомутанты мягкой пшеницы были использованы как непосредственно для создания сортов, так и включены в различные программы скрещиваний, что позволило получить ценный исходный материал для селекции.. В конкурсном сортоиспытании сорта Лютесценс 43-04 (мутантно-сортовой гибрид: (Лютесценс 4-94 х Мутант 536) х Терция) и Лютесценс 162-00 (межмутантный гибрид: Мутант 536 х Мутант 758) имели существенные преимущества по урожайности зерна над соответствующим по скороспелости стандартом, характеризовались устойчивостью к полеганию и поражению бурой ржавчиной (таблица 6).
Таблица 6 – Характеристика новых сортов яровой пшеницы
Сорт | Вегетационный период, сутки | Урожайность зерна, т /га | ||||||
2006 г. | 2007 г. | 2008 г. | Ср. | 2006г. | 2007 г. | 2008 г. | Ср. | |
Памяти Азиева, st | 74 | 77 | 71 | 74 | 3,19 | 2,73 | 2,73 | 2,88 |
Лютесценс 43-04 | 76 | 78 | 71 | 75 | 3,47 | 3,20 | 3,64 | 3,44 |
Лютесценс162-00 | 75 | 78 | 71 | 75 | 4,04 | 3,12 | 3,36 | 3,51 |
НСР0,05 | 0,12 | 0,50 | 0,18 | 0,27 |
В результате проведённых исследований были созданы сорта яровой мягкой пшеницы: Славянка Сибири, Светланка и Серебристая.
Славянка Сибири (Мутант 777). Сорт включён в Госреестр селекционных достижений России в 2002 году. Сорт среднеспелый, вегетационный период 78-83 дня. Высокоустойчив к полеганию и осыпанию. Высокоурожайный, средняя урожайность по пару составила 2,95 т/га, по зерновым – 2,34 т/га. Сорт отличается высоким качеством, особенно по содержанию белка и сырой клейковины, соответственно до 19,0 и 38,4%. Рекомендуется для лесостепной зоны при посеве по предшественникам: пар, однолетние травы, озимые, пропашные и зерновые культуры. Особый интерес представляет для хозяйств, специализирующихся по птицеводству, поскольку, благодаря повышенной питательности зерна, добавление его к фуражному в соотношении 1:1 позволяет получать соответствующий ГОСТу комбикорм. Включён в Госреестр селекционных достижений России в 2002 году (А. с.№ 000, патент № 000).
Светланка. Сорт выведен путём индивидуального отбора из гибридной популяции Омская 23 х Целинная 26. Разновидность лютесценс. Относится к среднеранней-среднеспелой группе спелости (74-78 суток). Устойчив к полеганию и осыпанию. Характеризуется высокой засухоустойчивостью, как на раннем этапе развития растений, так и в целом за вегетационный период. Средняя урожайность по пару составила 2,70 т/га (+0,64 т/га к стандарту), по зерновым – 2,43 т/га (+ 0,39 т/га). Имеет зерно высокого качества (белок 17,2%, клейковина 31,3%). Включён в Госреестр селекционных достижений России в2004 году (А. с. № 000, патент № 000). Включён в Госреестр Республики
Казахстан в 2006 году (А. с. № 000).
Серебристая. Сорт создан из мутантно-сортовой популяции (ОмСХИ 6 х Заволжская) х (Росинка х Мутант 717). Среднепозднего типа, период вегетации 75-89 суток. Средняя урожайность 2,65 т/га. Устойчив к пыльной головне, меньше стандарта поражается твёрдой головнёй. Сорт засухоустойчив, устойчив к полеганию. Включён в Госреестр селекционных достижений России в 2011 году (А. с. № 000, патент № 000).
Выводы
1. В повышении генотипической изменчивости и устойчивости мягкой пшеницы к биотическим и абиотическим факторам внешней среды в условиях Западной Сибири, а также в создании нового исходного материала для селекции культуры в регионе эффективно использовать химические мутагены.
2. Установлена зависимость эффекта действия химических мутагенов на мягкую пшеницу от вида мутагена, дозы и генотипических особенностей исходных сортов мягкой пшеницы. Выявлено ингибирующее действие на популяционные признаки всех изучаемых мутагенов, особенно НДММ, ДЭС, НДЭМ, ЭИ; стимулирующее же действие зависело от вида мутагена, дозы и генотипа сорта. Наибольшее снижение продуктивности М1 у всех сортов вызвали мутагены НДММ и ЭИ во всех изучаемых дозах, ДМС и НДЭМ в высоких дозах при различных сроках хранения обработанных семян. Отмечено стимулирующее действие на количественные признаки ДЭС и ингибирующее – ЭИ, ДМС, НДЭМ. Установлено увеличение размаха и величины фенотипической изменчивости признаков в мутантных популяциях, особенно при воздействии ЭИ, НДММ, ДМС, причём изменчивость признаков в М1 была выше, чем в М2.
3. Доля вклада в полевую всхожесть семян М1 генотипа сортов и мутагенов одинакова, но более весомое влияние оказывает не вид, а доза мутагена. Отмечена значительная доля влияния мутагенов и на выживаемость растений М1, также преобладало влияние дозы. В продуктивность растений М1 основной вклад вносил генотип сорта, доля вклада вида мутагена была значительно выше доли его дозы. При разных сроках хранения обработанных семян доля вклада в изменчивость продуктивности М1 вида и дозы мутагена была равной, срок хранения обработанных семян оказал незначительное влияние.
На проявление количественных признаков оказали влияние, как вид мутагена, так и его доза и поколение популяции мутантов. Доля вклада мутагена преобладала над долей дозы, особенно в М1. Значительным вклад дозы мутагена был в озернённость колоса, продуктивность колоса и растения, массу 1000 зёрен. Значительно было влияние на общую и продуктивную кустистость, массу зерна колоса в М1 взаимодействия факторов мутаген-доза.
4. Воздействием химических мутагенов у сортов яровой пшеницы получены различные макромутации: скороспелые, позднеспелые, морфологические, системные. Бóльшее количество мутаций получено при обработке химическими мутагенами ДАБ, НДЭМ, ДЭС, больше обладающих комплексом хозяйственно-ценных признаков скороспелых форм – НДММ и НДЭМ.
5. Установлено существенное влияние условий среды, как на проявление мутантных признаков, так и на характер их наследования. Мутации одних и тех же количественных признаков имели разную генетическую природу. Были получены рецессивные, доминантные и полудоминантные мутации количественных признаков. В зависимости от гидротермических условий наблюдалось ослабление, иногда затухание мутантных признаков, а также изменение характера их наследования. Мутации скороспелости были как доминантные (Мутант 576, Мутант 577, Мутант 579), так и рецессивные (у большинства мутантов).
Выявлено плейотропное действие мутантного гена у мутантов мягкой пшеницы. Признаки плейотропного комплекса, находящегося под контролем мутантного гена, изменяются в различных направлениях. Изменение комплекса количественных признаков может быть обусловлено мутированием одного гена или независимыми мутациями нескольких генов.
6. Отмечена зависимость величины и изменчивости количественных признаков у озимых и яровых сортов мягкой пшеницы от экологических факторов. Установлено благоприятное влияние новой генотипической среды на выраженность большинства мутантных признаков. Хемомутанты способствуют улучшению, как отдельных количественных признаков, так и продуктивности растений в целом в различных гибридных популяциях мягкой пшеницы.
Доля вклада генотипической среды и условий выращивания в количественные признаки мутантно-сортовых гибридов зависела от генотипа мутанта и признака: на проявление одних признаков основное влияние оказал генотип, других – условия среды, третьих – генотип, условия среды и взаимодействие факторов.
7. Наследование количественных признаков в мутантно-сортовых популяциях имеет различные типы – от сверхдоминирования до депрессии. На проявление мутантных признаков у мутантов мягкой пшеницы и их гибридов оказывают влияние условия выращивания и генотипическая среда. Идентификация генотипов более надёжна по признакам, контролируемым аддитивно действующими генами и проявляющими промежуточное наследование (высота растения, количество колосков в колосе). В разной степени выраженное доминирование по озернённости колоса, массе 1000 зёрен и массе зерна главного колоса затрудняет «узнавание» генотипа по фенотипу в благоприятных условиях среды, но в типичных для зоны условиях в отдельных комбинациях возможен отбор нужных генотипов по данным признакам. Коэффициенты наследуемости признаков продуктивности выше в
благоприятных по увлажнению и температурному режиму условиях выращивания, позволивших изучаемым популяциям реализовать генотипический потенциал наиболее полно.
8. Скрещивание мутантов с образцами, несущими гены устойчивости к
бурой ржавчине, позволило создать иммунные и высоко устойчивые к этому заболеванию образцы яровой мягкой пшеницы. Изучение у ржавчиноустойчивых форм показателей водного режима показало зависимость их от генотипа, фазы развития растений и гидротермических условий. Не обнаружено связи между интенсивностью транспирации и устойчивостью к ржавчине, наблюдалось как снижение, так и повышение признака у иммунных форм на инфекционном фоне. Выделены ржавчиноустойчивые формы с высокой водоудерживающей способностью, что указывает на их адаптивность к засушливым условиям.
9. Применение химических мутагенов позволило получить мутации совершенной яровости без значительных фенотипических перемен из сортов озимой пшеницы. На основе генетико-селекционного изучения гибридов и родительских форм яровой пшеницы установлено, что яровые фенотипические аналоги озимой пшеницы являются источниками генов для улучшения комплекса хозяйственно ценных признаков, связанных с продуктивностью и устойчивостью растений к полеганию.
10. С помощью экспериментального мутагенеза созданы адаптивные к условиям Западной Сибири формы озимой мягкой пшеницы, которые были использованы как непосредственно для создания новых сортов озимой пшеницы, так и в качестве доноров ряда признаков для улучшения озимой и яровой пшеницы. Успешный отбор адаптивных форм озимой пшеницы возможен в условиях вегетации, типичных для зоны.
11. У гибридов, полученных от скрещивания мутантов озимой пшеницы с сортами яровой, в первом поколении доминирует яровой образ жизни. В F2 наблюдалось расщепление популяций на яровые и озимые формы, в соотношении яровых и озимых растений по годам и комбинациям скрещиваний 15:1, 14:2, 13:3, 12:4.
Гибриды, полученные от скрещивания мутантов озимой пшеницы с сортами яровой, проявили гетерозис по отношению к яровому родителю и стандарту, как по продуктивности растения, так и по слагающим её элементам. Проявление гетерозиса носит дискретный характер и зависит как от комбинации скрещивания, так и от условий выращивания. В более благоприятных условиях роста и развития пшеницы наблюдался более высокий уровень гетерозиса. У большинства изученных гибридов во втором поколении гетерозис частично сохраняется. Фенотипическая изменчивость зависела от условий вегетации гибридов, в экстремальных условиях она повышалась. В благоприятных гидротермических условиях наблюдался более широкий размах трансгрессивной изменчивости. Отмечена зависимость проявления количественных признаков от условий выращивания, генотипа и генерации гибридных популяций. Отбор адаптивных форм необходимо проводить, начиная с F3, с последующим испытанием потомства.
12. Установлено наличие гена гибридного некроза Ne2ms у сорта Ильичёвка и мутантов Мироновской 808: Мутант 85 (сорт Омская озимая), Мутант 88, мутанта Ильичёвки: Мутант 33 (сорт Омская 3). В расщепляющихся популяциях F2, проявивших гибридный некроз в первом поколении, возможен отбор нормальных по фенотипу растений, превосходящих по продуктивности родительские сорта.
13. Отмечено изменение характера корреляционных взаимосвязей количественных признаков под влиянием мутагенов: мутационные воздействия нарушают сложившуюся и генетически обусловленную систему корреляций. Генотипическая среда существенно модифицирует характер взаимосвязи между признаками, что позволяет рекомендовать включение мутантов в скрещивания с целью создания нового ценного материала для селекции.
Анализ корреляционных плеяд показал их лабильность, обусловленную как генотипом, так и меняющимися условиями среды, причём влияние второго фактора проявляется сильнее. У мутантов и гибридов выявлена довольно устойчивая плеяда признаков: масса зерна - количество колосков - озернённость колоса.
14. Изучение комбинационной способности мутантов озимой и яровой пшеницы показало, что мутанты могут служить донорами как отдельных количественных признаков, так и результирующего показателя – урожайности зерна. Признаки в первой, второй и третьей генерации озимо-яровых гибридных популяций в большей степени контролировались действием аддитивных генов мутантов озимой пшеницы. Влияние взаимодействия генов родительских форм было значительно меньшее и в большой степени зависело от экологических факторов.
У гибридов мутантов яровой пшеницы независимо от поколения и условий года в генетическом контроле признаков преобладали эффекты ОКС, указывающие на доминирующее положение аддитивных эффектов генов, что позволяет вести отбор уже в ранних поколениях гибридных комбинаций.
15. Получены сорта и формы озимой и яровой мягкой пшеницы, адаптированные к условиям Западной Сибири. Использование мутантного материала позволило создать три сорта озимой пшеницы – Омская озимая, Сибирская нива и Омская 3. В Государственный реестр селекционных достижений России включены сорта яровой пшеницы мутантный сорт Славянка Сибири, межсортовой гибрид Светланка, мутантно-сортовой гибрид Серебристая. Создан новый перспективный материал, испытывающийся в различных селекционных питомниках лаборатории селекции пшеницы и озимого тритикале ФГБОУ ВПО ОмГАУ им. .
Практические рекомендации
1. С целью увеличения генотипической изменчивости мягкой пшеницы и создания адаптированных к условиям Западной Сибири форм озимой и яровой пшеницы рекомендуется использование химических мутагенов НДММ (дозы –0,05; 0,025; 0,012%), НДЭМ (дозы–0,02; 0,01%), и ЭИ (доза – 0,005%).
2.Отбор нужных биотипов следует проводить по признаку «количество колосков колоса», находящемуся в устойчивой коррелятивной зависимости с продуктивностью колоса и растения. Отборы высокопродуктивных адаптивных форм мягкой пшеницы необходимо проводить в годы, типичные по условиям вегетации для данной зоны.
3. Выделенные иммунные к бурой ржавчине (Г 566/01, Г 535/01, Г 620/01, Г 695/01, Г 697/01, Г 528/01, Г 581/02, Г 582/02) и высокоустойчивые (Г 583/01, Г 536/02 и Г 573/02) образцы и устойчивые к засухе формы (Г 566/01, Г 581/02) рекомендуется использовать для повышения адаптивного потенциала мягкой пшеницы.
4. На увеличение количественных признаков яровой пшеницы рекомендуется использовать мутанты озимой пшеницы сибирского генофонда. Мутантные сорта озимой пшеницы Сибирская нива и Омская 3 рекомендуется использовать в селекции озимой пшеницы, как доноров зимостойкости и хозяйственно-ценных признаков. В качестве комплексных улучшателей сортов яровой пшеницы предлагается использовать мутанты Мутант 562, Мутант 771, Мутант 772, Мутант 777, Мутант753.
5. При использовании в гибридизации озимой пшеницы сортов Ильичёвка, Омская озимая, Омская 3 и Мутанта 88 учитывать присутствие в их геноме доминантного гена гибридного некроза – Ne2ms.
Основные результаты исследования отражены в следующих публикациях:
Публикации в журналах, рекомендованных ВАК РФ
1. Кротова мутагены как фактор получения различных мутаций у яровой мягкой пшеницы /// Вестник Алтайского ГАУ, 2009. № 9 (59). С. 12-15.
2. Кротова оценки образцов яровой мягкой пшеницы, полученных с участием сортов и мутантов, в условиях юга Западной Сибири / , // Труды Кубанского ГАУ, 2009. № 6 (21). C. 58-61.
3. Поползухина признаков у хемомутантов яровой пшеницы и их гибридов / , // Известия СПбГАУ, 2009. № 17. С. 59-61.
4. Кротова скороспелых форм яровой мягкой пшеницы с помощью химических мутагенов/ // Вестник Алтайского ГАУ, 2010. №2 (64). С. 28-31.
5. Кротова способность мутантов и линий яровой пшеницы по основным элементам продуктивности/, // Вестник Алтайского ГАУ,2010. № 3 (65). C. 36-41.
6. Кротова мутагенных факторов и генотипической среды на характер корреляционных связей между признаками у яровой пшеницы / // Известия СПбГАУ, 2010. № 18. С. 29-31.
7. Кротова -генетическая оценка сортов и мутантов яровой мягкой пшеницы в условиях юга Западной Сибири / , // Вестник Саратовского ГАУ им. , 2010. № 04. C. 17-20.
8. Кротова зависимость основных хозяйственно-ценных признаков у мутантно-сортовых гибридов яровой пшеницы /, // Вестник Алтайского ГАУ, 2010. №4 (66). С. 16-20.
9. Роль химических мутагенов в получении адаптивных форм яровой мягкой пшеницы / //Известия СПбГАУ, 2010. № 21. С. 26-29.
10. Кротова -генетическая оценка мутантов яровой мягкой пшеницы /, // Известия ТСХА, 2010. № 5. С. 103-108.
11. Кротова химических и биологических мутагенов на взаимосвязи количественных признаков у мягкой пшеницы / , //Вестник Алтайского ГАУ, 2011. № 1. С. 45-48.
12. Кротова изучения влияния химических мутагенов на
яровую мягкую пшеницу / // Вестник Саратовского ГАУ им. , 2011. № 02. C. 18-23.
13. Кротова цитоплазматической наследственности на продуктивность колоса мутантно-сортовых гибридов / // Вестник Саратовского ГАУ им. , 2011. № 07. C. 9-11.
14. Создание адаптивных сортов яровой мягкой пшеницы для Западно-Сибирского региона /, , // Омский научный вестник, 2012. №1(108). C. 181-184.
Монографии
15. Леушкина -генетические аспекты адаптивности яровой мягкой пшеницы к условиям южной лесостепи Западной Сибири: монография / , , // Омск: Изд-во ФГОУ ВПО ОмГАУ, 20с.
16. Кротова генетического потенциала мутантов озимых форм в селекции мягкой пшеницы Западной Сибири: монография / , , /Омск: Изд-во ФГОУ ВПО ОмГАУ им. , 20с.
17.Кротова химических мутагенов на яровую мягкую пшеницу: монография/-LAMBERT Academic Publishing, 20с.
Авторские свидетельства, патенты
18. Авторское свидетельство № 000 РФ. Сорт яровой мягкой пшеницы Славянка Сибири /, , / СибНИИСХ. Заявка №9 дата приоритета 04.12.97 г., г.
19. Патент № 000 РФ. Сорт яровой мягкой пшеницы Славянка Сибири/ , , /СибНИИСХ. Заявка №9 дата приоритета 04.12.97 г., зарегистрирован 14.01.2003 г.
20. Авторское свидетельство № 000 РФ. Сорт яровой мягкой пшеницы Светланка/, , / СибНИИСХ. Заявка № 9 дата приоритета 10.12.2000 г., г.
21. Патент № 000 РФ. Сорт яровой мягкой пшеницы Светланка / , , А.Н. Ковтуненко, ., /СибНИИСХ. Заявка № 9 дата приоритета 10.12.2000 г., зарегистрирован 26.12.2003 г.
22. Авторское свидетельство № 000 Республика Казахстан. Сорт яровой мягкой пшеницы Светланка/ , , А.Н. Ковтуненко, , /СибНИИСХ. Заявка №0 дата приоритета 23.03.2002г., выдано 03.г.
23. Авторское свидетельство № 000 РФ. Сорт яровой мягкой пшеницы Серебристая/, , /СибНИИСХ. Заявка №9 дата приоритета 02.12.2008 г., г.
24. Патент № 000 РФ. Сорт яровой мягкой пшеницы Серебристая/ , , /СибНИИСХ. Заявка № 9 дата приоритета 02.12.2008 г., г.
Научные статьи и материалы
25. Кротова гибридного некроза у мутантно-сортовых гибридов / // Семеноводство и селекция сельскохозяйственных культур в Западной Сибири: Сб. науч. тр. / ВАСХНИЛ СО. СибНИИСХ. Новосибирск, 1990. С. 71-79.
26. Кротова развития гибридов, полученных от скрещивания мутантов озимых форм с сортами яровой пшеницы///Роль науки в интенсификации сельского хозяйства / Мат. конф. (г. Омск, 20 апреля 1989 г.). Ч.1. Растениеводство. Новосибирск, 1990. С. 52-55.
27. Кротова всходы–колошение и тип развития гибридов, полученных скрещиванием мутантов озимых форм с сортами яровой пшеницы/ , // Биология, селекция и генетика пшеницы и тритикале в Западной Сибири: Сб. науч. тр./ Омский СХИ. Омск, 1990. С. 45-50.
28. Белецкая наследования признаков продуктивности гибридами яровой пшеницы, созданными на мутантной основе / , //Биология, селекция и семеноводство полевых культур: Сб. науч. тр. / ОмСХИ. Омск, 1991. С. 10-16.
29. Рутц мутагенез в селекции яровой пшеницы / , // Селекция зерновых культур в Западной Сибири: Сб. науч. тр. /РАСХН. Сиб. отд-ние. СибНИИСХ. Новосибирск,1992. С.14-22.
30. Поползухина мутагенез в улучшении качества зерна яровой мягкой пшеницы/, //Биология, селекция и технологии возделывания сельскохозяйственных культур в Западной Сибири: Сб. науч. тр. / ОмГАУ. Омск, 1998. С. 54-58.
31. Храмцова комбинационной способности сортов и форм яровой пшеницы по массе зерна главного колоса в топкроссных скрещиваниях / , , // Биология, селекция и технологии возделывания сельскохозяйственных культур в Западной Сибири: Сб. науч. тр. / ОмГАУ. Омск, 1998. С. 39-43.
32. Храмцова комбинационной способности сортов и мутантных форм яровой пшеницы по озерненности главного колоса в топкроссных скрещиваниях / , , // Биологические особенности и приемы повышения продуктивности с./х. и местных культур. Сб. науч. тр. седьмой науч. конф. проф.-препод. состава и аспирантов агрономического факультета ОмГАУ (22 марта 2001 г.) Омск, ОмГАУ, 2002. С. 63-66.
33. Леушкина -биологическая характеристика устойчивых к бурой ржавчине образцов яровой пшеницы в условиях южной лесостепи Западной Сибири/, , //Новейшие направления развития аграрной науки в работах молодых учёных / Тр. 2 Междунар. науч.-практ. конф. молодых учёных. Новосибирск, 2006. С. 233-238.
34. Кротова мутационной изменчивости в селекции пшеницы///М-лы Междун. конферен. «Научное наследие – фундамент развития отечественного и мирового сельского хозяйства», 27-28 ноября 2007 г. М: ФГОУ ВПО РГАУ–МСХА им. , 2007. С. 78-79.
35. Храмцова комбинационной способности исходного материала для селекции яровой пшеницы/, // М-лы Междун. конферен. «Научное наследие – фундамент развития отечественного и мирового сельского хозяйства», 27-28 ноября 2007 г. М: ФГОУ ВПО РГАУ – МСХА им. , 2007. С. 127-128.
36. Кротова химических мутагенов на продуктивность растений М1 яровой мягкой пшеницы///М-лы Междун. научно-технич. Форума «Реализация Госуд. Программы развития с.-х. и регулирование рынков с.-х. продуктов, сырья и продовольствия: инновации, проблемы, перспективы (26-27 февраля 2009г.). Омск; Изд-во ФГОУ ВПО ОмГАУ, 2009. Ч.1. С. 214-217
37. Кротова действия химических мутагенов на яровую мягкую пшеницу///Экспериментальный мутагенез в биологии и сельском хозяйстве / Мат. Междунар. науч.-практ. конф.: Сб. науч. тр. Киров: Вятская ГСХА,2009. С. 58-61
38. Кротова наследования мутантных признаков у яровой мягкой пшеницы /, // Роль аграрной науки в инновационном развитии агропромышленного комплекса: Мат. Междунар. науч.-практ. конф., посвящ. 90-летию агрон. фак-та КазГАУ-Казань,2009.- С.100-103.
39. Кротова химических мутагенов на полевую всхожесть семян и выживаемость растений М1 яровой мягкой пшеницы/// Современные тенденции развития земледелия и защиты почв: Мат. Междунар. науч.-практ. конф. Улан-Удэ: изд-во БГСХА им. В.Р. Филиппова, 2009. С. 121-125.
40. Генетический потенциал мутантов в создании сортов яровой мягкой пшеницы для Западно – Сибирского региона/ , , // Экспериментальный мутагенез в биологии и сельском хозяйстве / Мат. Междунар. науч.-практ. конф.: Сб. науч. тр. Киров: Вятская ГСХА, 2009. С. 100-105.
41. Поползухина адаптивных сортов яровой мягкой пшеницы для Западно - Сибирского региона в условиях меняющегося климата/ , , // Климат, экология, сельское хозяйство Евразии. Иркутск, 2009. С. 63-68.
42. Кротова генотипа и условий среды на изменчивость признаков у сортов и мутантов яровой пшеницы/, //Аграрная наука — сельскому хозяйству: Сб. ст.: в 3 кн. /V Междунар. науч.-практ. конф. (17-18 марта 2010 г.). Барна0. Кн. 2. С. 300-302.
43. Кротова показателей водного режима у мутантов яровой мягкой пшеницы///Сибирская деревня: история, современное состояние, перспективы развития: Сб. науч. тр. в 3 частях. Омск, «Омскбланкиздат», 2010. Ч. III. С. 311-315.
44. Кротова экологических факторов и генотипической среды на вегетационный период и продуктивность колоса мутантов яровой пшеницы/ //Вавиловские чтения – 2010: Мат. Междунар. науч.-практ. конф. в 3 томах. Саратов: Изд-во КУБИК, 2010. Т.1. С. 42-44.
45. Кротова химических мутагенов на адаптивные свойства мягкой пшеницы/, // Актуальные вопросы экологии и природопользования: Мат. II Междунар. науч.-практ. конф. Ставрополь, 2011. С. 64-66.
46. Кротова химических мутагенов в повышении адаптивности яровой пшеницы/ , // Инновационные технологии создания и возделывания сельскохозяйственных
растений: Мат. Междунар. науч.-практ. конф./ под ред. Дужкина : Изд. КУБИК, 2011. С. 55-57.
47. Кротова адаптивности озимой пшеницы к условиям Западной Сибири с помощью химических мутагенов/ // Вавиловские чтения – 2011: Мат. Междунар. науч.-практ. конф. Саратов: Изд-во КУБиК, 2011. С. 38-40.
48. Кротова мутантного гена в различных условиях среды у яровой пшеницы/ //Актуальные научные вопросы: реальность и перспективы: Сб. науч. тр. по мат. Междунар. заоч. науч-практ. конф. 26 декабря 2011 г.: в 7 частях. Часть 3. Тамбов, 2012. С. 75-77.
49. Кротова корреляционных плеяд количественных признаков мутантов озимой пшеницы от экологических факторов/// Сибирская деревня: история, современное состояние, перспективы развития: Сб. науч. тр. в 3 частях. Омск: дом «Наука», 2012. Ч. III. С. 269-272.
50. Кротова наследственность в формировании количественных признаков у мутантно-сортовых гибридов яровой пшеницы/ // Научная жизнь, 2012. №3. С.63-70.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
