

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Результаты по величине полученных поправочных коэффициентов приведены в таблице № 5.
Таблица № 5. Поправочные коэффициенты для расчета при анализе многокомпонентных смесей с использованием системы дифференциальной детекции на основе штамма Comamonas testosteroni BS1310(pBS1010).
Тип анализируемой смеси | В присутствии катехола | В отсутствие катехола |
ТС – единственное сульфоароматическое соединение в смеси | 108% | 24% |
Присутствует несколько сульфоароматических соединений | 47% | 11% |
Из таблицы видно, что наиболее эффективно использование сенсора будет для определения общего количества сульфоароматических соединений в смеси, в которой катехол отсутствует или содержится в незначительном количестве.
В рамках данной работы было произведено сравнение характеристик клеток плазмидсодержащего штамма, выращенных на бензолсульфонате и толуолсульфонате. Их чувствительность к этим веществам достоверно не различалась (данные не представлены), что может служить косвенным подтверждением того, что деградация бензолсульфоната и п-толуолсульфоната осуществляется одним ферментом.
В заключение отметим, что в данной работе была создана модель сенсора, позволяющего определять концентрацию п-толуолсульфоната с нижним пределом детекции около 5 мкМ. Это обстоятельство, в сочетании с высокой селективностью анализа, делает возможным использование разработанной модели в качестве прототипа промышленного анализатора арилсульфонатов. Чувствительность сенсора к ацетату, этанолу и глюкозе не следует рассматривать в качестве ожидаемой помехи для анализа, поскольку данные вещества не встречаются в промышленных сточных водах.
В литературе отсутствуют сведения о работах по созданию микробного сенсора для определения сульфоароматических соединений. Имеются данные экспериментов по исследованию субстратной специфичности штаммов Pseudomonas rathonis T, Pseudomonas sp. 2T/1, P. aeruginosa 1S, P. putida K, Achromobacter eurydice TK [206]. В них было показано отсутствие чувствительности к сульфоароматическим соединениям. Таким образом, штамм C. testosteroni BS1310 (pBS1010) обладает достаточной специфичностью для определения сульфоароматики, что и определило его пригодность для детекции сульфоароматических соединений.
Другой путь оптимизации анализа может быть основан на свойстве клеток, состоящем в том, что одной из первых стадий деградации арилсульфонатов данной культурой является их десульфонирование с высвобождением сульфита. Таким образом, принципиально возможной схемой может быть формирование гибридного анализатора, включающего колоночный реактор с иммобилизованными клетками C. testosteroni BS1310 и биосенсор для детекции сульфита, аналог которого описан в работе [34]. Такой подход, хотя и связанный с некоторыми трудностями (необходимость оптимизации работы
реактора), позволил бы ограничить спектр детектируемых субстратов только сульфоароматическими соединениями.
4.2.4. Исследование работы сенсора на основе клеток Comamonas testosteroni в проточной системе
Целью данного раздела работы была проверка возможности использования разработанного сенсора для детекции ТС на основе плазмидсодержащего штамма Comamonas testosteroni в проточной системе. Данная постановка вопроса была связана с тем, что для детекции исследуемого вещества в реальных образцах сточных и природных вод зачастую необходимы приборы проточного типа, позволяющие производить измерения в непрерывном режиме.
Была сконструирована модель сенсора на основе клеток C. testosteroni для детекции ТС в проточной системе. Исследования проводили с использованием дозатора, позволяющего вводить пробы строго определенного объема (70 мкл). При иммобилизации на ДЭАЭ-целлюлозе штамм С. testosteroni, обладающий капсулой, вымывался из носителя, в отличие от штамма, лишенного капсулы. Исходя из этого, можно сделать предположение о том, что бескапсульный вариант в большей степени способен к адгезии на поверхностях.
Ранее исследование кюветной системы показало, что при отсутствии перемешивания в кювете концентрация кислорода уменьшается по сравнению с исходной концентрацией в перемешиваемой кювете до 75%.В проточной системе таких эффектов не наблюдается.
Были исследованы параметры работы системы в условиях протока без микроорганизмов. А затем эксперименты проводились в проточной системе с иммобилизованными в агаровом геле на поверхности электрода Кларка микроорганизмами. Наименьшая скорость, при которой было возможно проведение измерений, составляла 0.5 мл/ мин. Содержание кислорода при такой скорости составляло%, амплитуда ответа -% при концентрации ТС 10 мМ. Время прохождения пробы через рецепторную часть электрода при скорости 0.5 мл/ мин составляло 8.5 секунды. На более высоких скоростях протока амплитуда ответа была существенно ниже, а время отмывки короче. С учетом того, что время ответа электрода согласно паспортным данным составляет 90 с, оптимальным объемом пробы при данной скорости будет объем порядка 750 мкл (в 7 раз меньше объема стационарной кюветы). При таком объеме пробы можно будет получать ответы со значительной амплитудой.
Нижний предел детекции ТС штаммом C. testosteroni в проточной системе при скорости 0.5 мл/мин составил 5 мкМ, что сравнимо с чувствительностью в стационарной кювете. Но, учитывая короткое время контакта (8.5 с), чувствительность может оказаться выше, чем в стационарной кювете. Верхний предел детекции составил порядка 1 мМ, что также сопоставимо с данным показателем для стационарной системы. Но амплитуды ответов на высокие концентрации были существенно ниже по сравнению с кюветным методом. Это также можно объяснить коротким временем контакта вещества с электродом. На рис. 9 представлена калибровка по ТС в проточной системе на скорости 0.05 мл/мин с иммобилизованными на GF микроорганизмами. При данных условиях сенсор давал устойчивый ответ на концентрации толуолсульфоната 250 нМ, поскольку время контакта пробы с раствором составляло 85 с. Базовый уровень при этом имел вид сжатой синусоиды, шумы убирали с помощью программы "sensor". Скорость развертки на полярографическом анализаторе составляла см = 200 с).
Колебания базового уровня содержания кислорода в проточной системе при наличии микроорганизмов на электроде имели меньшую амплитуду, чем без микроорганизмов. Очевидно, это объясняется тем, что микроорганизмы поддерживают относительно стабильный уровень дыхания, и поэтому эффект от каждой новой порции раствора сглаживался.
Проверка субстратной специфичности в проточной системе дала те же результаты, что и в стационарной кювете. Получены высокие ответы на ацетат, этанол, глюкозу. Время, в течение которого сенсор на основе микроорганизмов, иммобилизованных на бумаге GF, проявлял стабильность показателей, составляло от нескольких часов до двух дней, т. е. столько же, сколько в стационарной кювете.
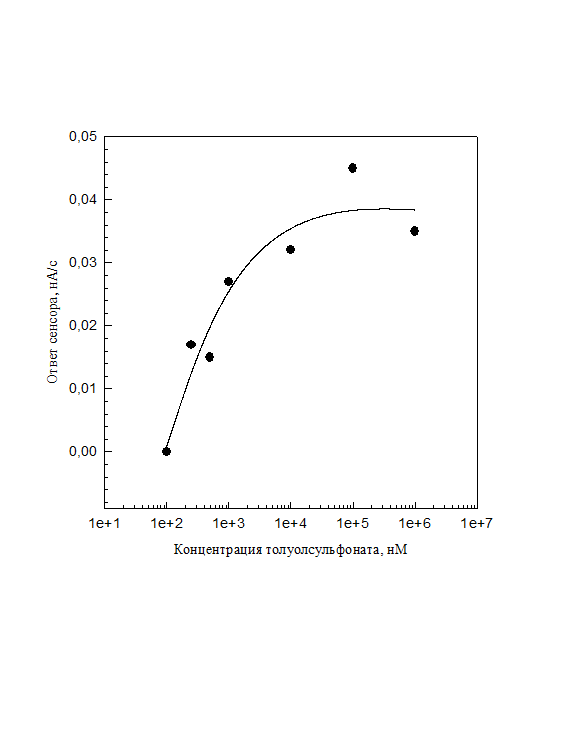
Рис. 11. Калибровка биосенсора на основе иммобилизованных в агаровый гель клеток C. testosteroni BS1310(pBS1010) в проточной системе при скорости протока 0.05 мл/мин.
4.3. Разработка микробного биосенсора для детекции фенола
4.3.1.Скрининг штаммов-деструкторов фенола
Поиск микроорганизмов, способных к деструкции фенола, производился среди штаммов, выделенных из нефтяных месторождений Западной Сибири сотрудниками Лаборатории биологии плазмид ИБФМ РАН.
Характерной особенностью бактериальных клеток, выделенных из почв нефтяных месторождений Западной Сибири, являлась их способность к росту на нефти и нефтепродуктах (дизельное топливо, мазут) в качестве единственного источника углерода и энергии. Из 300 культур микроорганизмов-деструкторов нефти к росту на моноциклических ароматических субстратах, в том числе на феноле, были способны 28 штаммов. Культуры существенно различались по скорости роста на феноле; для исследований были отобраны 9 штаммов, отличавшихся наибольшей скоростью роста. Отобранные микроорганизмы принадлежали к грамотрицательным бактериям и были использованы для формирования биорецепторов сенсора. Поскольку одной из основных характеристик сенсора является его избирательность по отношению к анализируемому веществу, представлялось целесообразным начать исследование с оценки субстратной специфичности штаммов.
Результаты оценки субстратной специфичности сенсоров на основе исследованных штаммов приведены на рис. 12. Для оценки субстратной специфичности использовали вещества различных классов - спирты, сахара, фенолы. Из приведенных на рисунке величин аналитических сигналов видно, что добавление фенола и катехола у всех исследованных штаммов активировало дыхание, что свидетельствует о потенциальной возможности их использования в рецепторных элементах.
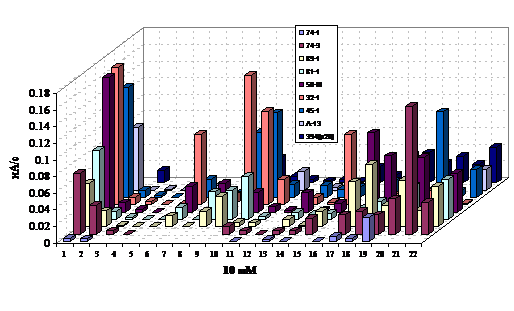
Рис. 12. Cубстратная специфичность исследованных штаммов. Все субстраты подавались в концентрации 10 мМ.
1-фенол, 2-этанол, 3-глицерин, 4-сорбит, 5-сорбоза, 6-ксилоза, 7-бутанол, 8-изопропанол, 9-глюкоза, 10-ксилит, 11-нафталин, 12-арабит, 13-катехол, 14-метанол, 15-пропанол, 16-изобутанол, 17-ацетат, 18-п-толуолсульфонат, 19-бензолсульфонат, 20-арсенит, 21-динитроортокрезол, 22-гранозан.
Штаммы:
1-394(р20), 2-А-13, 3-45-1, 4-32-I, 5-50-III, 6-81-I, 7-89-I, 8-74-III, 9-74-I.
Численные значения по субстратной специфичности фенолдеградирующих штаммов приведены в таблице № 4.
Таблица № 6. Субстратная специфичность исследованных фенолдеградирующих штаммов.
Штамм № п/п | 74-1 | 74-3 | 89-1 | 81-1 | 50-III | 32-1 | 45-1 | A-13 | 394(p20) |
1 | 0.0038 | 0.0731 | 0.052 | 0.0831 | 0.162 | 0.1654 | 0.1323 | 0.0754 | 0 |
2 | 0.0029 | 0.0346 | 0.019 | 0.0092 | 0.011 | 0.0092 | 0.0085 | 0 | 0.0146 |
3 | 0 | 0.0038 | 0.0015 | 0.0023 | 0.0031 | 0.0039 | 0.0023 | 0.0008 | 0 |
4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
5 | 0 | 0 | 0 | 0 | ингибир. | 0 | 0 | 0 | 0 |
6 | 0 | 0 | 0.0131 | 0.0146 | 0.031 | 0.085 | 0.022 | 0.0054 | 0 |
7 | 0 | 0 | 0 | 0 | ингибир. | 0 | 0 | 0 | 0 |
8 | 0 | 0 | 0.0185 | 0.0331 | 0.0342 | 0.0115 | 0.007 | 0.011 | 0 |
9 | 0 | 0 | 0.037 | 0.0346 | 0 | 0.1562 | 0.078 | 0.0077 | 0.03 |
10 | 0 | 0.0096 | 0.0054 | 0.0515 | 0.024 | 0.113 | 0.102 | 0.0054 | 0.0077 |
11 | 0 | 0.0038 | 0.0046 | 0.0038 | 0.0062 | 0.0031 | 0.0015 | 0.0023 | 0.0039 |
12 | 0 | 0 | 0 | 0 | 0.0023 | 0 | 0 | 0 | 0.0023 |
13 | 0.0019 | 0.0038 | 0.0085 | 0.0088 | 0.0231 | 0.0085 | 0.015 | 0.0031 | 0.0042 |
14 | 0 | 0.0038 | 0.0019 | 0.0062 | 0.0019 | 0.0031 | 0.01 | 0 | 0.0015 |
15 | 0 | 0.0192 | 0.0185 | 0.0077 | 0.01 | 0.0085 | 0.0123 | 0.0046 | 0.0192 |
16 | 0 | 0 | 0 | 0 | 0.0012 | 0 | 0 | 0 | 0.0092 |
17 | 0.0058 | 0.0231 | 0.054 | 0.0138 | 0.0954 | 0 | 0.019 | 0.0019 | 0.0331 |
18 | 0.0004 | 0.0027 | 0.075 | 0.0215 | 0.0677 | 0.0131 | 0.0154 | 0.0038 | 0.0362 |
19 | 0.0003 | 0.0023 | 0.026 | 0.0077 | 0.0038 | 0.0065 | 0.007 | 0.0015 | 0.0231 |
20 | 0 | 0.0042 | 0.056 | 0.04 | 0.0662 | 0.0169 | 0.1031 | 0.0046 | 0.032 |
21 | 0 | 0.0154 | 0.02 | 0.01 | 0.0092 | 0.0019 | 0.007 | 0 | 0.022 |
22 | 0 | 0.0039 | 0.049 | 0.0485 | 0.0462 | 0.0015 | 0.034 | 0.0254 | 0.0423 |
Помимо амплитуды сигнала на введение фенола, с целью подбора штамма, наиболее подходящего для его детекции, было рассчитано соотношение величины сигнала сенсора на фенол к общей сумме величин сигналов на остальные вещества. Назовем эту величину условно "Критерий селективности". Данные расчетов и амплитуды сигналов на фенол приведены в таблице № 5.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 |
