

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Таблица № 7. Величины сигналов сенсоров на фенол и отношение сигналов на фенол к общему мешающему сигналу.
Исследуемый штамм | Амплитуда сигнала на фенол (Сф) | Сумма сигналов, вызванных присутствием мешающих соединений (Сп) | Критерий селективности, Сф/Сп |
74-1 | 0.00038 | 0.0017323 | 0.22 |
74-3 | 0.00731 | 0.03865 | 0.19 |
89-1 | 0.0052 | 0.0408 | 0.127 |
81-1 | 0.00831 | 0.03133 | 0.265 |
50-III | 0.0162 | 0.04365 | 0.371 |
32-1 | 0.01654 | 0.04419 | 0.374 |
45-1 | 0.01323 | 0.044915 | 0.294 |
A-13 | 0.00754 | 0.007752 | 0.973 |
394(p20) | 0 | 0.02813 | 0 |
В таблице № 7 отображен расчет субстратной специфичности для плазмидного и бесплазмидного вариантов штамма 32-I по аналогии со штаммом C.testosteroni.
Как видно из таблицы, наибольшим значением критерия селективности обладал штамм А-13, второе по величине значение было у штамма 32-1. В то же время, сигналы у штамма А-13 имели низкую амплитуду по отношению ко всем веществам. Кроме того, для штамма А-13 не было показано наличие плазмиды биодеградации фенола. Больший интерес представлял штамм 32-I, обладающий плазмидой. Наличие плазмиды позволяло исследовать характеристики сенсоров на основе плазмидного и бесплазмидного вариантов для оценки возможности их использования в качестве дифференциальной пары для селективной детекции фенола (по аналогии с плазмидным и бесплазмидным штаммом C.testosteroni).
4.3.2. Характеристика штамма 32-I. (Субстратная специфичность)
Наибольшее внимание для дальнейшего исследования заслуживали параметры штамма 32-I, для которого характерным являлись наиболее высокие значения аналитических сигналов в ответ на введение фенола и катехола, превышавшие отклики на сахара в 5 – 7 раз; остальные штаммы характеризовались более низкой избирательностью по отношению к фенолу. Для всех штаммов была характерна широкая субстратная специфичность, изменение дыхания (сигналы сенсоров) в ответ на введение в измерительную кювету моносахаров (кроме глюкозы и ксилозы) было менее существенным (в 2 – 3 раза ниже), чем в ответ на добавление низкомолекулярных спиртов с неразветвленной цепью (этанол, пропанол, бутанол). Таким образом, при измерении концентрации фенола в средах сложного состава следует учитывать возможность искажения сигнала при наличии в среде низкомолекулярных спиртов с неразветвленной цепью.
Изучение генетического контроля деградации фенола показало, что, по крайней мере, у двух штаммов, 32-I и 83-IV, в трансформации этого соединения участвуют плазмиды размером около 100 тпн. Одним из доказательств плазмидного контроля деградации фенола является тот факт, что полученный нами бесплазмидный вариант штамма 32-I утрачивал способность к росту на феноле.
4.3.3. Характеристика сенсоров на основе плазмидсодержащего и бесплазмидного вариантов штамма 32-I
Штамм 32-I (плазмидный вариант) проявил высокую избирательность по отношению к фенолу, величина отклика сенсора на его основе в присутствии фенола существенно превышала величину отклика на другие использовавшиеся в эксперименте соединения. Этот штамм, несущий плазмиду биодеградации фенола, был исследован более детально как потенциальная основа рецептора биосенсора для определения фенола в водных растворах.
На рис. 13 представлен сравнительный портрет субстратной специфичности плазмидсодержащего и бесплазмидного вариантов штамма 32-I. Сенсор на основе бесплазмидного варианта был нечувствителен к фенолу; чувствительность к остальным субстратам была сравнима с таковой для сенсора на основе плазмидсодержащего варианта штамма 32-I.
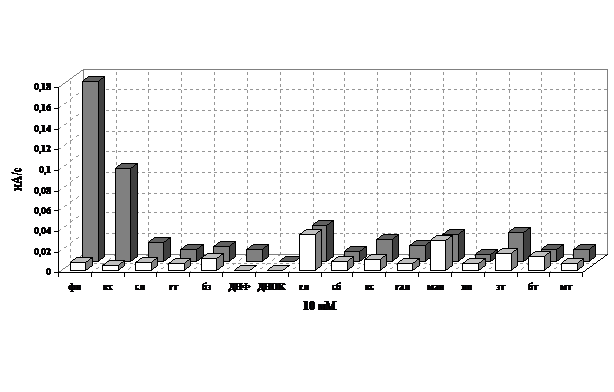
Рис. 13. Диаграмма субстратной специфичности сенсоров на основе плазмидного и бесплазмидного вариантов штамма 32-I. Субстраты вводились в концентрации 10 мМ.
Обозначение субстратов: фенол –1. катехол – 2, салицилат – 3, гентизат – 4, бензоат – 5, ДНФ – 6, ДНОК – 7, глюкоза – 8, сорбоза – 9, ксилоза – 10, галактоза – 11, манноза – 12, изопропанол – 13, этанол – 14, бутанол – 15, метанол – 16.
Таблица № 8. Анализ субстратной специфичности сенсоров на основе плазмидного и бесплазмидного варианта штамма 32-I.
Исследуемое вещество | Величина сигнала сенсора на основе бесплазмид-ного штамма | Величина сигнала сенсора на основе плаз-мидного штамма | Разница величин сигналов сенсоров | |
1 – фенол | 0.0082 | 0.175 | 0.167 | |
2 – катехол | 0.0047 | 0.091 | 0.086 | |
3 – салицилат | 0.0082 | 0.019 | 0.0105 | |
4 – гентизат | 0.007 | 0.012 | 0.0047 | |
5- бензоат | 0.012 | 0.014 | 0.002 | |
6 – ДНФ | 0 | 0.012 | 0.012 | |
7 – ДНОК | 0 | 0 | 0 | |
8 - глюкоза алкилалкилбензолсульфонат алкилбензолсульфонат алкилбензолсульфонат | 0.035 | 0.035 | 0 | |
9 – сорбоза | 0.009 | 0.009 | 0 | |
10 – ксилоза | 0.0105 | 0.021 | 0.0105 | |
11 – галактоза | 0.007 | 0.015 | 0.008 | |
12 – манноза | 0.029 | 0.026 | -0.003 | |
13 – изопропанол | 0.007 | 0.007 | 0 | |
14 – этанол | 0.016 | 0.028 | 0.012 | |
15 – бутанол | 0.014 | 0.012 | -0.002 | |
16 – метанол | 0.007 | 0.012 | 0.005 | |
Суммарная разница | 0.1457 | |||
Разница величин сигналов на целевое вещество | 0.167 |
|
Погрешность (добавочный сигнал) при определении фенола в присутствии всех мешающих веществ составляет 87,2%, в присутствии всех веществ, кроме катехола – 35,7%. При осуществлении дифференциальной детекции необходимо будет нормировать показания с учетом этих расчетов. При интегральном определении всех ароматических соединений, содержащихся в пробе, погрешность на мешающие вещества составляет 12.7%. Поэтому использовать данный сенсор для интегрального определения фенолов в реальных образцах можно при анализе стоков, содержащих преимущественно фенольные соединения. В этом случае погрешность, вызванная присутствием остальных классов соединений, будет незначительной, поскольку величина 12.7% получена для ситуации, при которой все соединения (и целевые, и мешающие), находятся в среде в одинаковой концентрации.
Таким образом, селективность сенсора на основе штамма 32-I сопоставима с селективностью сенсора на основе C.testosteroni (11% и 12%, соответственно). Данные по величине погрешности для дифференциальной пары на основе штамма 32-I представлены в таблице № 9.
Таблица № 9. Величины погрешности для дифференциальной пары на основе штамма 32-I при различных условиях исследования.
Тип анализируемой смеси | В присутствии катехола | В отсутствие катехола |
Фенол – единственное ароматическое соединение в смеси | 87.2% | 35.7% |
Присутствует несколько ароматических соединений | 12.7% |
Калибровочная зависимость плазмидсодержащего штамма 32-I, полученная для фенола, представлена на рис.14. Диапазон детекции составлял 5 – 1000 мкМ фенола. Более высокие концентрации фенола вызывали ингибирование дыхания клеток. Этот эффект, по-видимому, носит достаточно общий для микробных клеток характер, поскольку аналогичные результаты были получены для амперометрического биосенсора на основе Pseudomonas putida [142].
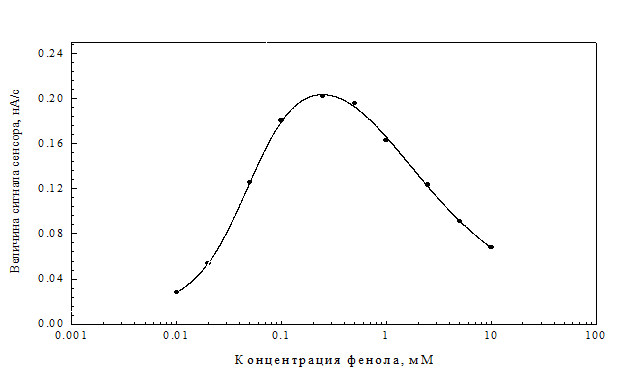
Рис. 14. Калибровочная кривая сенсора на основе иммобилизованных клеток плазмидсодержащего варианта штамма 32-I. Исследуемое вещество - фенол.
Следует отметить, что нижний предел детекции фенола для сенсора на основе штамма 32-I был сопоставим с аналогичным параметром для моделей сенсоров, описанных в литературе. Так, нижний предел детекции для сенсора на основе Trichosporon cutaneum составил 2 мкМ [55], для сенсора на основе Rhodococcus его величина составила 4 мкМ [165].
На рис. 15 изображена зависимость ответов сенсора от величины рН, которая показывает, что оптимальным для функционирования клеток являлось значение рН 7.4. Температурная зависимость сенсора показана на рис. 15. Максимальные значения аналитических сигналов были получены при температуре 35°С.
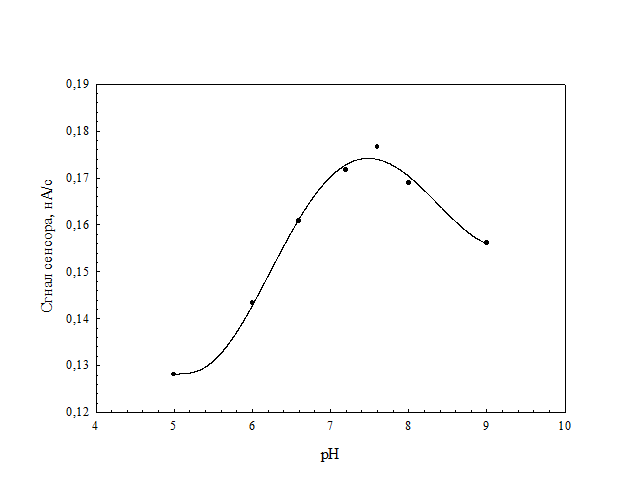
Рис. 15. Зависимость величины ответа сенсора на основе иммобилизованных клеток плазмидсодержащего варианта штамма 32-I от величины рН. Среда измерения – калий-фосфатный буфер 30 мМ. Концентрация фенола – 25 мкМ.
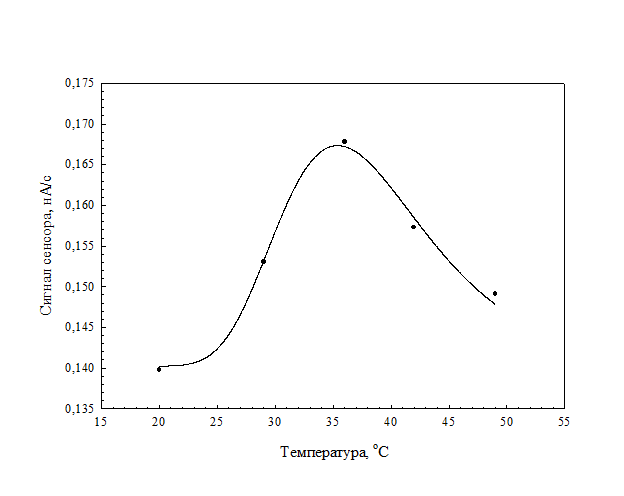
Рис. 16. Кривая зависимости величины сигнала сенсора от температуры.
Полученные данные по рН и температурному оптимуму для микроорганизмов позволяют произвести сравнение с аналогичными параметрами для ферментов, проявляющих фенолоксидазную активность Так, известно, что для b-тирозиназы оптимальными является рН 9.0 и температура 30оС, для фенол-2-монооксигеназы – рН 7.6 и температура 22оС, для полифенолоксидазы – рН 7.0 и температура 25оС, для лакказы – рН 7.0 и температура 37оС [207]. Таким образом, можно допустить, что для штамма 32 - I характерна лакказная активность.
Стабильность сигналов сенсора изображена на рис. 16. Время жизни составляло 30 часов. Стабильные ответы (без потери активности) наблюдали на протяжении 8 часов, что несколько меньше, чем у описанных в литературе сенсоров. В частности, для сенсора на основе Rhodococcus [165] время жизни составляло 21 день, время жизни сенсора на основе P. putida DSM-548, описанного в работе [142], также составляло 21 день.
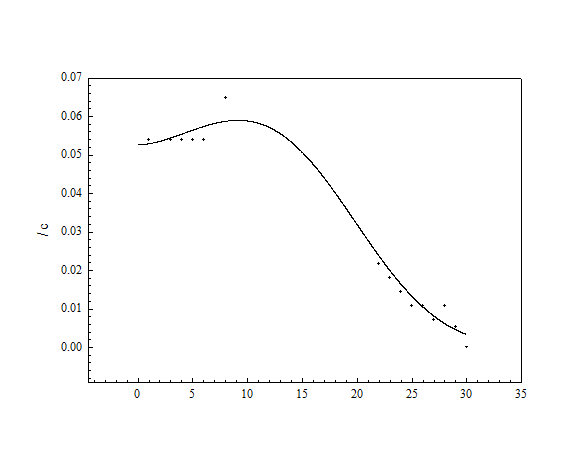
Рис. 17. Кривая стабильности сенсора на основе плазмидсодержащего варианта штамма 32-I. В качестве контрольного был использован раствор фенола в концентрации 25 мкМ.
Причиной малого времени жизни сенсора предположительно могли быть два фактора: вымывание клеток из носителя и их инактивация или гибель. С целью проверки предположения о возможности вымывания клеток из состава биорецептора было проведено исследование стабильности сенсора при иммобилизации клеток в агаровый гель, поскольку включение в гель обеспечивает более прочное закрепление клеток, чем адсорбция. Кроме того, известно, что при микробной деградации токсичных соединений иммобилизация в агаровый гель оказывает защитное действие на клетки микроорганизмов [148]. В нашем случае стабильность клеток, иммобилизованных в агаровый гель, была сравнима со стабильностью клеток, адсорбированных на бумаге. Следовательно, возможной причиной малого времени жизни сенсора является снижение ферментативной активности клеток или их гибель. Нашли, что активность сенсора снижалась пропорционально по отношению ко всем веществам, что позволяет сделать вывод об инактивации клеток в целом, а не ферментных систем, отвечающих за окисление фенола. Можно предположить, что иные методы иммобилизации, позволяющие обеспечить оптимальные условия для клеток, приведут к увеличению стабильности сенсора.
Поскольку чувствительность сенсоров на основе плазмидсодержащего и бесплазмидного варианта штамма 32-I ко всем веществам, кроме фенола и катехола, была примерно одинакова, мы предположили, что в данной ситуации имеется возможность использования пары сенсоров на основе плазмидсодержащего и бесплазмидного штамма в качестве основы для дифференциальной детекции фенола в средах сложного состава. В этом случае разность сигналов сенсоров позволит получить более точную оценку содержания фенола в средах сложного состава.
4.4. Возможные пути решения практических задач с применением биосенсорного подхода
Применение созданных лабораторных макетов биосенсоров на основе штамма-деструктора ТС и штамма 32-I для анализа сточных вод нефтеперерабатывающего производства (г. Лисичанск, предприятие Лисичанскнефтеоргсинтез) показало, что специфические сигналы сенсора отсутствуют и основным эффектом при контакте сенсора со стоками является ингибирование клеточного дыхания. Наблюдаемый эффект объяснили присутствием токсичных компонентов стоков. На основе лабораторных анализов по основным компонентам стоков можно предположить, что ингибирование было обусловлено присутствием значительного количества токсичной органики (химическое потребление кислорода - ХПК - 300 мг/л и более) и сульфидов (более 20 мг/л). Это послужило основанием для выполнения серии тестов, основанных на полученном ранее опыте по изучению деградации ТС в колоночном реакторе, содержащем C. testosteroni BS1310 (pBS1010) и направленном на исследование возможности использования для определения токсичных соединений в сточных водах микроорганизмов активного ила, применяемых при их очистке. В данном случае в колоночном реакторе вместо бактериальных клеток применили активный ил водоочистных сооружений нефтеперерабатывающего производства, который, по сути, является более адаптированным биоматериалом для подобных анализов. Был также сформирован сенсор для детекции фенола с использованием активного ила в качестве рецепторного элемента. Сенсор обладал широчайшей субстратной специфичностью и характеризовался почти полным отсутствием избирательности. Только при введении нитроароматических соединений (2, 4-ДНФ, пикриновая кислота, мочевина) наблюдались несколько более высокие амплитуды сигналов сенсора. Для всех остальных тестированных веществ (спирты, сахара, органические кислоты) амплитуды сигналов сенсора были практически одинаковыми. Для сенсора была построена калибровочная зависимость по фенолу с использованием стандартных растворов (рис. 18). Нижний предел детекции составил 5 мкМ (как и для сенсора на основе штамма 32-I), наибольшая амплитуда сигналов наблюдалась при концентрации фенола 200 мкМ. При наличии высоких концентраций фенола наблюдался эффект субстратного ингибирования.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 |
