

Семикратное введение исследованных препаратов при аллоксановом диабете, в отличие от однократного режима их применения, в подавляющем большинстве случаев вызывало уменьшение содержания продуктов ПОЛ в крови больных крыс. В первую очередь это касалось реамберина, все дозы которого при этом режиме применения достоверно снижали уровень изопропанолрастворимых ДК. Коме того, минимальная доза данного ЛС обусловила достоверное снижение содержания гептанрастворимых КД и СТ, а его средняя дозировка привела к убыли изопропанолрастворимых КД и СТ. Полученные данные на первый взгляд диссонируют с результатами оценки влияния реамберина на липидную пероксидацию in vitro и свидетельствуют о непрямом (опосредованном) влиянии реамберина на систему «ПОЛ – АОЗ» in vivo. α-ЛК, также как реамберин обладавшая проксидантным действием in vitro, в условиях курсового 7-дневного применения тоже вызывала достоверное уменьшение уровня продуктов ПОЛ. Это касалась изопропанолрастворимых ДК, содержание которых значимо уменьшилось при использовании относительно высоких доз α-ЛК (ЭСТД и 2 ЭСТД), а также изопропанолрастворимых КД и СТ, уровень которых снизился при курсовом введении минимальной и максимальной доз. Особого обсуждения заслуживает изменение уровня продуктов ПОЛ при 7-кратном применении производных 3-оксипиридина (эмоксипина и мексидола). Невзирая на свое однозначно антиоксидантное действие in vitro, эмоксипин при 7-кратном режиме применения вызвал существенное увеличение уровня гептанрастворимых ДК при использовании относительно низких доз (½ ЭСТД и ЭСТД). 7-кратное введение максимальной дозы эмоксипина вызвало уменьшение содержания гептанрастворимых КД и СТ в крови больных крыс. Повторное введение эмоксипина на протяжении 7 дней оказало разнонаправленное действие на уровель изопропанолкастворимых ДК в зависимости от использованной дозировки. Минимальная доза эмоксипина снижала содержание обсуждаемой категории липопероксидов в крови больных животных, в то время как средняя доза, наоборот увеличивала уровень этой категории продуктов в сыворотке крови. Мексидол, который также как реамберин является сукцинатсодержащим ЛС, при 7-кратном введении максимальной дозы, подобно реамберину, снижал уровень изопропанолрастворимых ДК в крови крыс с аллоксановым диабетом. Вместе с тем, повторное введение мексидола в относительно низкой дозе (ЭСТД), как и использование аналогичной дозы эмоксипина, оказывало прооксидантное действие по критерию накопления циркулирующих изопропанолрастворимых ДК.
Более продолжительное (14-дневное) введение всех изученных ЛС вызывало однонаправленное нарастание уровня циркулирующих продуктов ПОЛ. В случае эмоксипина это проявилось достоверным увеличением содержания гептанрастворимых ДК и изопропанолрастворимых КД и СТ при использовании минимальной дозы этого ЛС, а также нарастанием уровня гептанрастворимых КД и СТ в случае применения его средней дозировки. Реамберин во всем диапазоне изученных доз вызывал достоверный прирост содержания изопропанолрастворимых ДК, т. е. оказывал действие, диаметрально противоположное соответствующему эффекту при его 7-дневном применении. Кроме того, 14-кратное введение реамберина в максимальной дозе достоверно увеличивало содержание гептанрастворимых КД и СТ. Последствия 14-дневного применения α-ЛК весьма напоминали результаты применения реамберина и заключались в значимом нарастании уровня изопропанолрастворимых ДК во всем диапазоне изученных доз. При использовании в средней терапевтической дозе α-ЛК дополнительно вызывала значимое нарастание уровня гептанрастворимых КД и СТ. Следует подчеркнуть, что ПОЛ-модулирующее действие реамберина и α-ЛК при 14-дневном режиме их применения хорошо согласуется с данными о их прооксидантном потенциале in vitro. Мексидол, одновременно являющийся производным 3-оксипиридина и янтарной кислоты, оказывал такое же как эмоксипин, но менее выраженное действие. 14-кратное введение мексидола в минимальной дозе вызывало достоверное нарастание изопропанолрастворимых КД и СТ, а в средней дозе дополнительно увеличивало уровень гептанрастворимых ДК.
Полученные данные не позволяют расценивать влияние исследованных ЛС на систему «ПОЛ – АОЗ» в качестве главного, и тем более, единственного молекулярного механизма их влияния на метаболические процессы. Следует подчеркнуть, что производные 3-ОП и ЯК ни в одном из режимов применения не оказали значимого влияния на показатели токоферолемии и содержание церулоплазмина, являющихся узловыми компонентами антиоксидантной защиты плазмы крови. Единственное исключение составил препарат сравнения α-ЛК, значимо снижавший концентрацию циркулирующего церулоплазмина с 44,17±3,95 до 33,94±4,14 мг/дл (p = 0,043) при однократном введении максимальной дозы в острый период аллоксанового диабета. По-видимому, в целом фазное влияние изученных препаратов на липопероксидацию при аллоксановом диабете является вторичным, сложно опосредованным феноменом, имеющим косвенное отношение к терапевтической эффективности исследованных средств при СД. Понимание природы обсуждаемой динамики продуктов ПОЛ под действием изученных ЛС возможно лишь на основе интегрированного анализа их эффектов на циркулирующий и тканевой пул продуктов свободнорадикального окисления липидов.
Влияние изученных ЛС на церебральные процессы ПОЛ существенно различалось в зависимости от локализации отделов мозга. Наиболее яркие эффекты были отмечены на уровне корковых структур и существенно зависели от кратности введения препаратов. Однократное введение изученных ЛС не оказывало никакого влияния на содержание липофусцин-позитивных нейронов в поле Par1 первичной соматосенсорной коры. При 7-кратном применении лишь мексидол, используемый в средней дозировке, вызвал значимое нарастание доли липофусцин-позитивных нейронов в I-III слоях изученной структуры неокортекса c 17,89±1,01% у крыс с аллоксановым диабетом, получавшим только инсулинотерапию, до 38,45±7,87% (p = 0,011). Этот факт соответствует ранее отмеченному приросту уровня изопропанолрастворимых ДК в крови больных животных и подтверждает положение о прооксидантном действии 7-дневного использования ЭСТД мексидола.
Последствия двухнедельного введения изученных ЛС соответствовали их антигипоксической активности и влиянию на процессы ПОЛ in vitro. Производные 3-оксипиридина (эмоксипин и мексидол) во всем диапазоне изученных доз в 1,7 - 8 раз снижали количество липофусцин-содержащих нейронов в поверхностных слоях поля Par1 (рисунок 3). Двухнедельное применение всех доз реамберина и α-ЛК, напротив, вызывало значимое нарастание доли липофусцин-позитивных нейронов в поверхностных слоях неокортекса. Аналогичная направленность действия изученных ЛС отмечена и на уровне глубоких слоев первичной соматосенсорной коры.
На уровне древней коры изученные препараты в подавляющем большинстве случаев оказывали однонаправленное антиоксидантное действие. При однократном введении наиболее ярко этот эффект проявился у эмоксипина, минимальная и максимальная дозы которого вызывали достоверное уменьшение содержания липофусцин-позитивных нейронов в поле СА1 гиппокампа больных животных с 19,15±1,33% до 14,57±0,97% (p = 0,022) и 12,33±0,85% (p = 0,004) соответственно. При обсуждаемом режиме введения мексидол оказывал такое же действие лишь в средней дозировке, снижая долю липофусцин-позитивных нейронов с 19,77±1,99% до 13,62±1,39% (p = 0,022). Ни одна из доз α-ЛК не повлияла на содержание липофусцина в нейронах поля СА1 аммонова рога при однократном введении. Отдельного внимания заслуживают эффекты реамберина, который, невзирая на свою прооксидантную активность, при однократном введении в минимальной дозе практически двукратно (с 18,28±2,13% до 9,92±1,69%, p = 0,032) снижал долю липофусцин-позитивных нейронов поля СА1 гиппокампа у крыс с аллоксановым диабетом. Важно подчеркнуть, что эффект аналогичной направленности, но несколько меньшей выраженности сохранялся после 7-дневного введения реамберина в минимальной дозе, когда содержание нейронов с липофусцином уменьшилось с 20,38±3,96% до 12,22±2,97% (p = 0,022). 7-кратное введение минимальной дозы α-ЛК, также, как реамберин, обладавшей прооксидантным действием in vitro, в два раза (с 20,38±3,96% до 10,32±2,19%, p = 0,022) снижало долю липофусцин-позитивных нейронов в поле СА1. При этом средняя доза α-ЛК оказывала противоположное действие, слабо, но статистически значимо (с 20,38±3,96% до 22,72±4,17%, p = 0,028) увеличивая содержание липофусцин-позитивных нейронов в изученной структуре древней коры. 7-кратное введение мексидола не оказало никакого влияния на содержание липофусцин-позитивных нейронов в поле СА1. Эмоксипин, в отличие от мексидола, сохранял антиоксидантную направленность действия на уровне гиппокампа крыс с аллоксановым диабетом при введении в минимальной дозе, что проявилось уменьшением доли липофусцинпозитивных нейронов с 19,83±4,90% до 13,77±1,31% (p = 0,043).
Важно подчеркнуть, что 14-дневное применение всех изученных средств вызывало однонаправленное значимое снижение процентного содержания липофусцин-содержащих нейронов поля СА1 во всем диапазоне использованных доз (рисунок 3). По-видимому, отмеченный эффект может быть связан с функциональной кумуляцией антиоксидантного эффекта для производных 3-оксипиридина (эмоксипина и мексидола) и ко-антиоксидантного эффекта для реамберина и α-ЛК.
На диэнцефальном уровне изученные ЛС влияли на процентное содержание липофусцин-позитивных нейронов только при курсовом введении. Все препараты оказали качественно однонаправленное прооксидантное действие, выраженность которого существенно зависела от влияния исследованных препаратов на липидную пероксидацию in vitro.
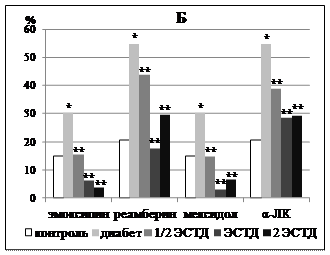
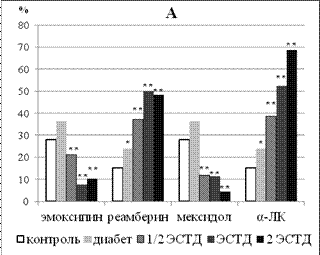
Рисунок 3 – Влияние 14 - кратного введения α-липоевой кислоты, производных 3-оксипиридина и янтарной кислоты на процентную долю липофусцин-содержащих нейронов в поверхностных слоях поля Par 1 первичной соматосенсорной коры (А) и поля СА1 гипокампа (Б) крыс с аллоксановым диабетом (* - p < 0,05 по сравнению с интактным контролем, ** - p < 0,05 по сравнению с группой животных с аллоксановым диабетом здесь и далее в рисунках 4 - 6)
ЛС с прооксидантной активностью (реамберин и α-ЛК) увеличивали долю липофусцин-позитивных нейронов уже при 7-кратном введении минимальной дозы с 26,32±2,47% у крыс с аллоксановым диабетом, получавших только инсулинотерапию, до 42,94±5,30% (реамберин, p = 0,039) и до 42,33±3,00% (α-ЛК, p = 0,003). При 14-кратном введении соответствующий эффект наблюдался во всем диапазоне исследованных доз реамберина и α-ЛК. Реамберин увеличивал долю нейронов, содержащих липофусцин, в 1,1 - 1,2 раза (р = 0,006-0,027), α-ЛК – в 1,3 - 1,5 раза (р = 0,,036). Производные 3-оксипиридина вызывали аналогичный эффект только при двухнедельном применении отдельных доз. Средняя доза эмоксипина повышала долю липофусцин-содержащих нейронов с 26,91±0,89% до 33,73±2,78% (р = 0,035), а средняя и максимальная дозы мексидола – до 38,22±1,86% (р = 0,004) и 36,16±1,77% (р = 0,006) соответственно.
Установленные эффекты производных 3-ОП и ЯК в отношении динамики содержания липофусцин-позитивных нейронов в изученных структурах головного мозга можно трактовать лишь в сопоставлении с сопутствующими изменениями клеточного состава церебральных структур и сдвигами этологического статуса крыс с аллоксановым диабетом. Изученные ЛС в целом оказывали нейропротекторное действие у крыс с аллоксановым диабетом. Эта закономерность весьма ярко проявилась в отношении количества нейронов первичной соматосенсорной коры больных животных в условиях курсового применения изученных ЛС. При этом препараты, обладающие антигипоксическим действием и антиоксидантной активностью in vitro (эмоксипин и мексидол), характеризовались нарастанием выраженности нейропротекторного действия по мере увеличения длительности применения. В то же время, реамберин и α-ЛК, оказавшие прооксидантное действие in vitro, наоборот, демонстрировали наибольшую выраженность протекторного эффекта при 7-дневной схеме применения с последующим уменьшением соответствующего действия к 14-му дню курса. Как видно (рисунок 4А), эмоксипин и мексидол при 7-кратной схеме введения оказали нейропротекторный эффект в поверхностных слоях соматосенсорной коры всего в двух дозах. Это касается относительно высоких доз (ЭСТД и 2 ЭСТД) для эмоксипина и относительно низких доз (½ ЭСТД и ЭСТД) для мексидола. При увеличении длительности введения до 14 дней нейропротекторный эффект производных 3-оксипиридина в отношении нейронов поверхностных слоев неокортекса наблюдался во всем диапазоне исследованных доз (рисунок 4Б). При этом максимальная доза эмоксипина дополнительно повышала число нейронов глубоких слоев неокортекса с 268,06±34,79 на 1 мм2 у крыс с аллоксановым диабетом, получавших только инсулинотерапию, до 391,00±10,73 на 1 мм2 (p = 0,007). Максимальная и минимальная дозировки мексидола при 14-кратном введении также увеличивали число нейроцитов в IV-VI слоях поля Par1 до 381,85±22,07 на 1 мм2 (p = 0,034) и 413,13±33,30 на 1 мм2 (p = 0,021) соответственно.
Реамберин эффективно предотвращал убыль нейронов в поверхностных слоях неокортекса при 7-кратном введении всех изученных доз крысам с аллоксановым диабетом (рисунок 4А). Аналогичный эффект данного ЛС в отношении глубоких слоев соматосенсорной коры наблюдался только при введении минимальной и максимальной дозировок, повышавших число нейронов с 323,84±24,26 до 389,74±50,82 на 1 мм2 (p = 0,013) и 414,18±29,86 на 1 мм2 (p = 0,046) соответственно. α-ЛК увеличивала содержание неокортикальных нейронов в поверхностных слоях при семикратном применении относительно высоких доз (ЭСТД и 2ЭСТД). Такой же эффект в глубоких слоях коры развивался при 7-дневном применении максимальной дозы α-ЛК, когда число нейронов возрастало с 323,84±24,26 до 456,67±38,05 на 1 мм2 (p = 0,036). Увеличение длительности применения реамберина и α-ЛК до двух недель приводило к ослаблению их нейропротекторного действия. У реамберина этот эффект в отношении нейронов поверхностных слоев неокортекса сохранялся только для 2 ЭСТД, у α-ЛК - лишь для ЭСТД (рисунок 4Б) и не проявлялся на уровне глубоких слоев новой коры.
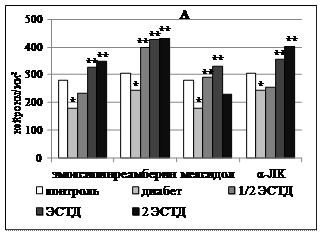
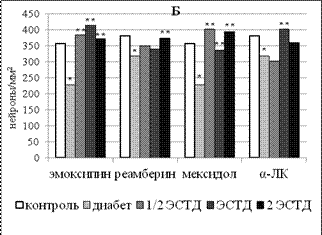
Рисунок 4 – Влияние 7 - кратного (А) и 14 - кратного (Б) введения α-липоевой кислоты, производных 3-оксипиридина и янтарной кислоты на количество нейронов поверхностных слоев поля Par 1 первичной соматосенсорной коры крыс с аллоксановым диабетом
Отдельного внимания заслуживает действие изученных ЛС на динамику другой категории терминально дифференцированных клеток нейроэктодермального происхождения (олигодендроцитов) в первичной соматосенсорной коре крыс с аллоксановым диабетом. При однократном режиме применения реамберин был единственным из изученных ЛС, оказавшим влияние на данный параметр. Это проявилось значимым снижением олигодендроцитов в поверхностных слоях поля Par1 при применении 2 ЭСТД реамберина с 104,55±9,87 до 73,98±2,23 на 1 мм2 (p = 0,007). В отличие от однократного, 7-кратное введение реамберина не оказало никакого действия на число олигодендроцитов неокортекса. 14-дневное применение ЭСТД этого ЛС вызвало значимый прирост числа олигодендроцитов как в поверхностных слоях поля Par1 (с 145,70±10,20 до 186,57±7,72 на 1 мм2, р = 0,007), так и в глубоких (с 163,33±14,26 до 214,25±5,39 на 1 мм2, р = 0,012). Аналогичное (двухфазное) действие на динамику числа олигодендроцитов оказала α-ЛК, которая значимо снижала анализируемый показатель в поверхностных слоях неокортекса при 7-кратном введении в средней дозировке (с 139,01±29,48 до 67,19±5,32 на 1 мм2, p = 0,038) и значимо, в 1,25 – 1,43 раза (р = 0,005 – 0,024), увеличивала число олигодендроцитов в глубоких слоях новой коры при 14-кратном применении всех изученных доз крысам с аллоксановым диабетом. При 7-кратном применении эмоксипина в поверхностных слоях соматосенсорной коры число олигодендроцитов уменьшалось с 118,74±30,55 до 62,31±8,32 на 1 мм2 (p = 0,009) и до 76,18±6,48 на 1 мм2 (p = 0,049) при использовании ½ ЭСТД и ЭСТД этого ЛС соответственно. В глубоких слоях неокортекса эти же дозы препарата снижали количество олигодендроцитов с 131,54±38,84 до 72,54±3,09 на 1 мм2 (p = 0,028) и до 62,39±7,63 на 1 мм2 (p = 0,027) соответственно. При 14-кратном курсовом применении эффект эмоксипина в отношении олигодендроцитов претерпевал инверсию в поверхностных слоях неокортекса, где ½ ЭСТД и ЭСТД препарата повышали число олигодендроцитов с 138,54±3,79 на 1 мм2 у крыс с аллоксановым диабетом, получавших только инсулинотерапию, до 155,20±14,27 на 1 мм2 (р = 0,022) и 171,81±10,37 на 1 мм2 (р = 0,021) соответственно. Характер влияния эмоксипина на число олигодендроцитов в глубоких слоях неокортекса не претерпел существенной динамики в сравнении с результатами 7-кратного введения этого ЛС. Мексидол оказывал влияние на олигодендроциты поля Par1 больных крыс лишь при двухнедельном введении максимальной дозы, которая вызвала нарастание их содержания в глубоких слоях неокортекса с 190,39±21,44 до 195,87±7,42 на 1 мм2 (р = 0,032).
Наиболее распространенная клеточная популяция ЦНС (астроциты) продемонстрировала иную динамику сдвигов под действием изученных ЛС, эффекты которых реализовались только при курсовом применении. В подавляющем большинстве случаев производные 3-ОП и ЯК, так же как и α-ЛК, способствовали увеличению числа астроцитов неокортекса у крыс с аллоксановым диабетом. 7-кратное введение эмоксипина в дозах ЭСТД и 2 ЭСТД привело к увеличению числа астроцитов в поверхностных слоях соматосенсорной коры с 69,01±6,40 до 109,04±8,26 на 1 мм2 (p = 0,014) и до 137,92±27,26 на 1 мм2 (p = 0,01) соответственно. Аналогичный эффект той же локализации наблюдался при 14-кратном применении минимальной и максимальной доз эмоксипина (рисунок 5А). В то же время, двухнедельное введение средней дозы эмоксипина снижало число астроцитов в IV-VI слоях поля Par1 с 225,15±10,96 до 201,12±7,23 на 1 мм2 (p = 0,021). Мексидол не влиял на число неокортикальных астроцитов при 7-кратном применении, но также как эмоксипин, увеличивал число этих клеток в поверхностных слоях неокортекса при 14-дневном введении минимальной и максимальной дозы. В отличие от эмоксипина, двухнедельное применение максимальной дозы мексидола дополнительно увеличивало число астроцитов в глубоких слоях соматосенсорной коры с 225,15±10,96 до 279,95±15,68 на 1 мм2 (p = 0,012). 7-кратное введение относительно высоких доз реамберина вызывало нарастание числа астроцитов во всех слоях неокортекса. Под действием ЭСТД этого ЛС количество астроцитов увеличилось в поверхностных слоях поля Par1 с 111,13±13,75 до 199,42±24,81 на 1 мм2 (p = 0,015), а при введении 2 ЭСТД – до 188,48±27,67 на 1 мм2 (p = 0,019). В глубоких слоях поля Par1 число астроцитов возросло с 135,16±14,84 до 202,74±23,15 на 1 мм2 (p = 0,021) и 205,19±29,55 на 1 мм2 (p = 0,019) соответственно. При 14-кратном применении аналогичный эффект наблюдался под действием всех изученных доз реамберина в глубоких слоях неокортекса, где количество астроцитов повысилось в 1,3 - 1,4 раза (р = 0,,034) в сравнении с больными животными, получавшими только инсулинотерапию. Семидневное применение α-ЛК в средней дозе также увеличивало содержание астроцитов в поверхностных слоях неокортекса с 111,13±13,75 до 171,12±25,03 на 1 мм2 (p = 0,035) и в глубоких слоях соматосенсорной коры – с 135,16±14,84 до 263,07±44,31 на 1 мм2 (p = 0,019). При 14-дневном применении эффект аналогичной направленности наблюдался только в поверхностных слоях новой коры под действием относительно низких доз α-ЛК - ½ ЭСТД и ЭСТД (рисунок 5А).
Влияние изученных ЛС на динамику глиоцитов мезенхимального происхождения (микроглиоцитов) у крыс с аллоксановым диабетом характеризовалось относительно меньшей выраженностью в сравнении с изменениями остальных клеточных популяций. При однократном режиме введения лишь реамберин увеличивал число неокортикальных микроглиоцитов в глубоких слоях при использовании минимальной дозы с 44,20±5,82 до 63,69±7,25 на 1 мм2 (p = 0,047). При 7-кратной схеме введения только эмоксипин в средней дозе уменьшал содержание микроглиоцитов в поверхностных слоях неокортекса с 42,96±7,67 до 17,94±3,98 на 1 мм2 (p = 0,05) и α-ЛК в максимальной дозе оказывала аналогичный эффект в глубоких слоях, снижая их число с 62,21±14,80 до 29,08±1,98 на 1 мм2 (p = 0,00дневное применение всех изученных средств оказало однонаправленное действие на содержание микроглиоцитов в поле Par1 (рисунок 5Б). Наибольшая дозовая распространенность этого действия отмечена у ЛС с прооксидантной активностью (реамберина и α-ЛК). Все дозы α-ЛК и относительно низкие дозы реамберина (½ ЭСТД и ЭСТД) вызывали нарастание числа клеток микроглии в глубоких слоях соматосенсорной коры. При этом средние дозировки реамберина и α-ЛК увеличивали также число микроглиоциов в поверхностных слоях неокортекса с 99,26±11,14 до 129,89±6,40 на 1 мм2 (p = 0,043) и 159,03±14,12 на 1 мм2 (p = 0,005) соответственно. Средства с антиоксидантной активностью, эмоксипин и мексидол, повышали число микроглиоцитов лишь в единичных дозах. Это касается 14-дневного применения ЭСТД эмоксипина, вызывавшего нарастание числа микроглиоцитов в поверхностных слоях поля Par1 с 120,69±10,99 до 131,81±10,66 на 1 мм2 (р = 0,032) и 2 ЭСТД эмоксипина, оказывавшего аналогичный эффект в глубоких слоях неокортекса (рисунок 5Б). Мексидол повышал содержание клеток микроглии только в глубоких слоях соматосенсорной коры при использовании максимальной дозы (рисунок 5Б).
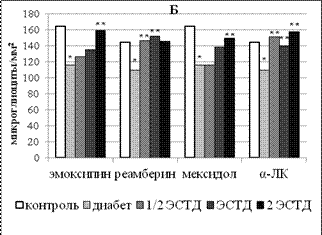
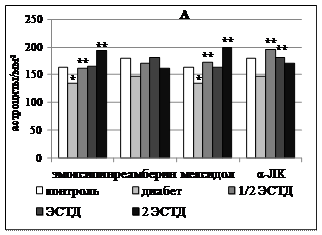
Рисунок 5 – Влияние 14 - кратного введения α-липоевой кислоты, производных 3-оксипиридина и янтарной кислоты на количество астроцитов поверхностных слоев (А) и микроглиоцитов глубоких слоев поля Par 1 (Б) первичной соматосенсорной коры крыс с аллоксановым диабетом
Особого внимания заслуживает влияние изученных ЛС на динамику основных клеточных популяций поля СА1 гиппокампа, роль которого в регуляции аффективного статуса и когнитивных функций общеизвестна. Все изученные препараты оказывали значимое влияние на показатели содержания нейронов в этой структуре аммонова рога уже при однократном применении. В первую очередь это касалось процентного содержания пирамидных нейронов поля СА1, которое значимо нарастало при однократном применении максимальных доз эмоксипина (с 14,38±1,45% до 18,27±1,28%, p = 0,037) и реамберина (с 16,16±3,01% до 28,76±2,24%, р = 0,028), средней дозы α-ЛК (с 13,05±1,86% до 16,97±2,99%, p = 0,048) и минимальной дозы мексидола (с 13,05±1,86% до 17,99±1,34%, p = 0,037), который дополнительно увеличивал долю корзинчатых нейронов с 11,97±1,38% до 18,71± 1,13% (p = 0,005). Реамберин оказался единственным ЛС, вызвавшим достоверное нарастание абсолютного количества корзинчатых нейронов в поле СА1 крыс с аллоксановым диабетом при использовании минимальной дозы с 213,73±15,15 до 387,27±92,31 на 1 мм2 (p = 0,028).
В дальнейшем, при курсовом применении исследованных ЛС, их влияние на клеточные популяции гиппокампа, так же как и на уровне первичной соматосенсорной коры, существенно зависело от анти - или прооксидантной активности in vitro. Минимальная доза реамберина снижала общее число нейроцитов поля СА1 с 2541,67±87,16 до 2121,20±290,47 на 1 мм2 (p = 0,013) и одновременно уменьшала процентное содержание корзинчатых клеток с 20,66±3,47% до 18,60±0,91% (p = 0,013) при 7-кратном курсовом применении. Средства с антиоксидантной активностью (эмоксипин и мексидол), наоборот, способствовали нарастанию числа нейронов гиппокампа в результате повторного 7-кратного введения. Данный режим применения эмоксипина был связан с увеличением как процентного содержания корзинчатых нейронов (с 14,28±0,83% до 20,82±1,75%, p = 0,028), так и их абсолютного количества (с 227,77±12,21 до 462,23±101,94 на 1 мм2, p = 0,048) при использовании максимальной дозы. Одновременно повышалось и общее число нейронов гиппокампа с 1980,36±63,52 до 2627,91±182,91 на 1 мм2 (p = 0,042). 7-кратное введение минимальной дозы мексидола привело к достоверному приросту абсолютного количества пирамидных нейронов (с 250,75±15,10 до 376,73±70,54 на 1 мм2, p = 0,028) и общего число нейронов (с 1980,36±63,52 до 2270,52±397,95 на 1 мм2, p = 0,015) в изученной структуре аммонова рога. Средняя доза мексидола также вызывала нарастание абсолютного числа пирамидных нейронов поля СА1 с 250,75±15,10 до 363,92±73,91 на 1 мм2 (p = 0,008) при недельном курсовом применении у крыс с аллоксановым диабетом.
Результаты 14-дневного введения исследованных препаратов также существенно зависели от их влияния на липидную пероксидацию in vitro. При этом реамберин и α-ЛК вызывали снижение содержания пирамидных нейронов поля СА1 древней коры. При двухнедельном применении всех дозировок α-ЛК абсолютное количество этой категории нейронов уменьшалось в 1,37 - 1,64 раза (р = 0,013 – 0,039), относительное – в 1,53 - 1,62 раза (р = 0,,013), а при введении средней дозы реамберина – в 1,54 раза (р = 0,021) и 1,42 раза (р = 0,043) соответственно. Следует заметить, что минимальная доза реамберина вызывала нарастание относительного содержания корзинчатых клеток с 19,34±0,83% до 21,07±2,21% (р = 0,019). Производные 3-ОП (эмоксипин и мексидол) оказали влияние совершенно иной направленности на содержание нейронов древней коры при аллоксановом диабете. 14-кратное курсовое применение минимальной дозы эмоксипина привело к значимому нарастанию относительного (с 21,31±1,34% до 25,09±2,22%, p = 0,037) и абсолютного (с 320,56±31,56 до 431,31±41,96 на 1 мм2, p = 0,049) количества корзинчатых нейронов поля СА1, а также повышению общего числа нейронов (с 1548,32±58,78 до 1882,89±20,89 на 1 мм2, p = 0,003). При увеличении дозировок отмеченные эффекты претерпевали инверсию, что проявилось достоверным снижением процентной доли пирамидных (с 31,62±2,90% до 20,60±2,14%, p = 0,028) и корзинчатых (с 21,31±1,34% до 18,32±0,37%, p = 0,045) нейронов при использовании средней дозы, а также процентного содержания пирамидных нейронов при использовании максимальной дозы (с 31,62±2,90% до 25,62±1,57%, p = 0,030). Мексидол, уступавший по антиоксидантной активности эмоксипину, при 14-дневном применении минимальной дозы вызвал лишь нарастание процентного содержания корзинчатых нейроцитов (с 21,31±1,34% до 25,05±1,49%, p = 0,037).
Влияние исследованных препаратов на содержание глиальных клеток при однократном режиме применения не зависело от их антиоксидантного потенциала. Данный режим применения выявил наличие существенного глиопротекторного эффекта только у двух препаратов: эмоксипина и реамберина. Однократное введение относительно высоких дозировок (ЭСТД и 2 ЭСТД) эмоксипина крысам с аллоксановым диабетом привело к значимому нарастанию олигодендроцитов в поле СА1 гиппокампа больных животных с 89,07±0,69 до 119,87±14,02 на 1 мм2 (p = 0,046) и до 166,67±19,46 на 1 мм2 (p = 0,046) соответственно. Аналогичный режим применения средней и максимальной дозировок реамберина сопровождался увеличением числа олигодендроцитов поля СА1 гиппокампа с 91,17±2,48 до 123,37±7,46 на 1 мм2 (p = 0,014) и до 117,00±4,78 на 1 мм2 (p = 0,004) соответственно. Кроме того, в минимальной дозе реамберин вызвал более чем двукратное нарастание содержания микроглиоцитов в обсуждаемой структуре древней коры (с 36,21±3,48 до 75,71±11,77 на 1 мм2, p = 0,009). Недельное курсовое применение изученных ЛС привело к нарастанию числа палеокортикальных олигодендроцитов только для средств с антиоксидантной активностью и отмечалось при использовании всех дозировок эмоксипина (в 1,59 - 2,69 раза, р = 0,,024) и мексидола (в 1,63 - 2,21 раза, р = 0,,009). α-ЛК, обладавшая наибольшим прооксидантным действием, вызвала уменьшение числа олигодендроцитов гиппокампа при использовании средней дозы с 115,84±13,52 до 74,71±7,78 на 1 мм2 (p = 0,030). Реамберин не оказал никакого влияния на содержание олигодендроцитов в древней коре при 7-кратном режиме применения, но одновременно вызвал достоверное нарастание числа астроцитов при использовании относительно низких доз: с 148,59±22,91 до 206,20±12,71 на 1 мм2 при введении ½ ЭСТД (p = 0,046) и до 240,37±13,89 на 1 мм2 при введении ЭСТД (p = 0,009). Такой же эффект в отношении клеток астроглии поля СА1 оказало 7-кратное введение максимальной дозы α-ЛК (увеличение числа астроцитов с 148,59±22,91 до 274,26±32,84 на 1 мм2, р = 0,026), средней дозы эмоксипина (с 109,51±10,21 до 255,39±24,86 на 1 мм2, р = 0,025) и минимальной дозировки мексидола (с 109,51±10,21 до 165,49±12,29 на 1 мм2, р = 0,025). Сукцинатсодержащие ЛС (реамберин и мексидол) вызвали уменьшение числа клеток микроглии в поле СА1 при 7-дневном применении относительно высоких доз реамберина (с 55,29±5,74 до 46,32±2,97 на 1 мм2 при введении ЭСТД, p = 0,040 и до 48,26±0,67 на 1 мм2 при введении 2 ЭСТД, p = 0,040), а также максимальной дозировки мексидола (с 64,38±1,87 до 52,68±2,94 на 1 мм2, p = 0,027). При этом следует подчеркнуть, что минимальная доза мексидола, наоборот, повышала число микроглиоцитов до 83,33±21,88 на 1 мм2 (р = 0,024), так же как и количество других глиальных клеток (астроцитов и олигодендроцитов), в изученной структуре аммонова рога.
Результаты 14-дневного применения исследованных ЛС в отношении числа астроцитов существенно зависели от наличия сукцината в составе изученных препаратов. Сукцинатсодержащие ЛС при 14-кратном применении минимальных доз вызывали значимое нарастание числа астроцитов в поле СА1 крыс с аллоксановым диабетом: реамберин – с 305,88±16,35 до 372,91±19,72 на 1 мм2 (p = 0,027), мексидол – с 321,29±19,96 до 381,28±16,50 на 1 мм2 (p = 0,036). Средние дозы эмоксипина оказали противоположный эффект, снизив число этой категории глиоцитов у крыс с аллоксановым диабетом с 321,29±19,96 до 299,72±7,68 на 1 мм2 (p = 0,032). α-ЛК не влияла на число палеокортикальных астроцитов при двухнедельном режиме применения. Количество олигодендроцитов и клеток микроглии нарастало под действием всех изученных ЛС при 14-кратном применении. Данный эффект наиболее ярко проявился при введении средства с наибольшей антиоксидантной активностью (эмоксипина), который вызвал прирост числа как олигодендроцитов (в 1,17 - 1,26 раза, р = 0,,021), так и микроглиоцитов (в 1,26 - 1,49 раза, р = 0,,016) во всех изученных дозах. Другое производное 3-оксипиридина (мексидол) вызвало нарастание олигодендроцитов с 200,51±10,67 до 256,64±14,45 на 1 мм2 (p = 0,015) и до 265,16±11,32 на 1 мм2 (p = 0,002) при использовании минимальной и максимальной доз соответственно, а также увеличение количества микроглиоцитов поля СА1 с 140,33±9,71 до 216,73±16,33 на 1 мм2 (р = 0,005) и до 189,68±8,58 на 1 мм2 (р = 0,022) при введении соответственно ЭСТД и 2 ЭСТД данного препарата крысам с аллоксановым диабетом. Относительно низкие дозы α-ЛК вызвали нарастание числа олигодендроцитов в аммоновом роге с 207,68±17,62 до 249,36±10,80 на 1 мм2 при применении ½ ЭСТД (р = 0,022) и до 252,97±5,24 на 1 мм2 при использовании ЭСТД (р = 0,049). Введение средней и максимальной доз этого ЛС привело к аналогичным изменениям числа микроглиоцитов палеокортекса больных животных, содержание которых повысилось с 158,12±12,32 до 211,08±3,79 на 1 мм2 (р = 0,015) и до 195,38±9,49 на 1 мм2 (р = 0,043) соответственно. Наименьшая «распространенность» соответствующего дозового эффекта была отмечена у реамберина, который при 14-кратном введении крысам с аллоксановым диабетом вызвал нарастание числа олигодендроцитов в средней дозе (с 207,68±17,62 до 272,94±16,35 на 1 мм2, р = 0,021) и микроглиоцитов в минимальной дозе (с 158,12±12,32 до 227,16±7,00 на 1 мм2, р = 0,005).
Обобщая данные о влиянии изученных ЛС на структуры палеокортекса, можно предположить их способность модулировать процессы пролиферации и дифференцировки плюрипотентных стволовых клеток древней коры, которая содержит значительную часть пула стволовых нервных клеток (Lee E., Son H. Adult hippocampal neurogenesis and related neurotrophic factors // BMB Rep. 2009. Vol. 42, № 5. Р. 239-244.). Высокая вероятность такого действия изученных препаратов связана с относительно слабовыраженными изменениями числа зрелых клеток нейроэктодермального происхождения (прежде всего нейронов) в изученной структуре аммонова рога в динамике аллоксанового диабета.
Не исключено, что действие исследованных ЛС на клеточный состав ПВЯ у крыс с аллоксановым диабетом тоже может быть в определенной степени связано с мобилизацией пролиферативного потенциала стволовых нервных клеток. Такая возможность иллюстрируется достоверным нарастанием числа нейронов ПВЯ с 371,36±44,17 до 444,13±16,01 на 1 мм2, p = 0,042) при однократном введении малой дозы эмоксипина. Наиболее выраженные эффекты изученных ЛС на диэнцефальном уровне были отмечены при 7-дневном применении. Важно подчеркнуть, что эти эффекты были однонаправленными, позитивными и не зависели от антиоксидантной и антигипоксической активности изученных ЛС. Недельное применение эмоксипина и реамберина вызвало значимое нарастание числа нейронов ПВЯ крыс с аллоксановым диабетом при использовании всех дозировок этих ЛС (рисунок 6А). α-ЛК оказала такое же действие при введении относительно высоких доз. При 14-кратном применении мексидол вызвал аналогичный эффект в единственной (минимальной) дозировке, повышая число нейронов ПВЯ с 532,48±15,62 до 626,59±41,31 на 1 мм2 (p = 0,034). Такой же режим введения α-ЛК привел к нарастанию числа нейронов ПВЯ с 529,96±25,30 до 631,17±41,49 на 1 мм2 (p=0,048). Двухнедельное применение эмоксипина и реамберина не оказало никакого влияния на содержание нейронов в ПВЯ при аллоксановом диабете.
Число глиальных клеток ПВЯ также изменилось уже при однократном применении изученных ЛС. При этом в максимальной дозе реамберин снижал число клеток астроглии с 278,69±18,91 до 223,41±14,34 на 1 мм2 (p = 0,032), а α-ЛК, напротив, увеличивала их количество с 188,54±7,78 до 231,03±11,85 на 1 мм2 (p = 0,016). Однократное введение средней дозы эмоксипина уменьшало число олигодендроцитов с 294,04±15,20 до 224,95±16,27 на 1 мм2 (p = 0,014). 7-ми кратное введение максимальной дозы α-ЛК, как и однократное, приводило к увеличению содержания астроцитов в ПВЯ больных животных с 246,30±19,07 до 367,14±31,53 на 1 мм2 (p = 0,004). Мексидол при недельном применении в максимальной дозе снижал число олигодендроцитов (с 420,89±131,80 до 211,28±8,79 на 1 мм2, p = 0,009) и микроглиоцитов (с 154,21±57,30 до 60,92±7,13 на 1 мм2, p = 0,040) в ПВЯ. 7-кратное введение эмоксипина и реамберина существенно не изменило глиальный состав изученного ядра гипоталамуса. Наиболее выраженный глиопротекторный эффект изученных ЛС наблюдался при их двухнедельном применении. При этом производные 3-оксипиридина (эмоксипин и мексидол), обладавшие антиоксидантной активностью in vitro, существенно превосходили средства с прооксидантной активностью (реамберин и α-ЛК) по «дозовой распространенности» влияния на глиальные популяции ПВЯ больных крыс. Двухнедельное введение эмоксипина и мексидола во всем диапазоне изученных доз вызвало достоверное нарастание числа астроцитов (рисунок 6Б) и олигодендроцитов (рисунок 6В) у крыс с аллоксановым диабетом. На этом фоне лишь минимальная доза эмоксипина вызывал значимое уменьшение количества клеток микроглии в ПВЯ (рисунок 6Г). Повышение числа астроцитов наблюдалось под действием не только максимальной, но и минимальной дозы α-ЛК (рисунок 6Б). Следует подчеркнуть, что 14-дневное курсовое применение относительно низких доз α-ЛК (½ ЭСТД и ЭСТД) дополнительно увеличивало число олигодендроцитов и микроглиоцитов ПВЯ крыс с аллоксановым диабетом (рисунок 6 В, Г).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
