

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
2.1 Физиологические свойства, определяющие функцию
глиальных клеток в полевом механизме сознания
Ряд морфологических характеристик глии и ее физиологические свойства предопределяют основную функцию глиальных клеток в составе полевого механизма сознания. Рассмотрим основные из них.
1. В ЦНС мембраны клеток разделены щелями в 15-20 нм и больше, однако между глиальными клетками очень часто образуются щели – "щелевые контакты" шириной 2 нм. Количество таких контактов в ЦНС возрастает в областях повышенного количества синаптических контактов между нейронами и их повышенной метаболической и функциональной активностью. Функциональная значимость этого явления не была выявлена. Предполагалось, что щелевые контакты имеют пониженное электрическое сопротивление, что облегчает электротоническое распространение тока ионов между глиальными клетками и, следовательно, выравнивание их концентрации в соседних клетках. Это могло бы означать синхронизацию неких процессов в глиальных клетках, обусловленную активностью нейронов. Как и другие проблемы, связанные с функцией глиальных клеток (например, с их чрезвычайной избыточностью), значение такой синхронизации было непонятно и оставалось неизвестным до возникновения концепции полевого механизма сознания и необходимости определения структур головного мозга, участвующих в его функционировании.
2. В ЦНС мембранный потенциал Ем глиальных клеток достигает высоких значений (до 100-110 и более мВ) и отличается относительной стабильностью – клетка не возбуждается при электрической стимуляции токами, вызывающими деполяризацию мембран нейронов. Однако она чрезвычайно чувствительна к наружной концентрации ионов калия [К+]н: мембранный потенциал Ем глиальной клетки изменяется на значительную величину при небольшом повышении концентрации [К+]н. Это обуславливает возникновение ее реакции на возбуждение близлежащей нервной клетки или нервного волокна, при котором ионы калия выходят наружу. Из-за малого объема жидкости в межклеточных щелях концентрация [К+]н в них может достичь значительных величин, что влечет за собой повышение концентрации К+ в близлежащих глиальных клетках и приводят к снижению мембранного потенциала Ем – деполяризации их мембран. Через щелевые контакты К+ могут проникнуть в соседние глиальные клетки, находящиеся на большем расстоянии от нервной клетки, и таким образом вокруг нервной клетки образуется оболочка из глиальных клеток-сателлитов с одинаково деполяризованными мембранами.
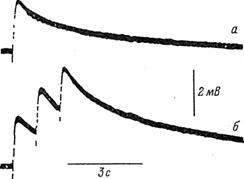
Нервный импульс длительностью в 1 мс приводит к деполяризации мембранного потенциала глиальной клетки Ем продолжительностью в несколько секунд [19]. По расчетам, увеличение [К+]н на
1 ммоль/литр в межклеточной щели шириной 20 нм вызывает деполяризацию мембраны глиальной клетки на 7,6 мВ ([1] с. 86). В экспериментах с препаратами различных биологических объектов деполяризация глиальных мембран длилась от нескольких секунд до нескольких десятков секунд. При частоте раздражения 1/с наблюдается суммация; при частоте 5/с и более эффекты от отдельных импульсов сливаются. При значении [К+]н = 20 ммоль/литр деполяризация может достичь 48 мВ. Длительность деполяризации (плато) может составлять 60 и более секунд.
На рис. 2.1 приведены результаты внутриклеточной регистрации изменения мембранных потенциалов глиальных клеток, возникавших при прохождении по волокну нервных импульсов. На рис. 2. 1(а и б) видно, что один проходящий по волокну нервный импульс вызывает деполяризацию мембраны глиальной клетки более чем на 2 мВ. Три следующих по аксону импульса с частотой 1/с приводят к суммации ее ответа: Еm мембраны возрастает на 4,5 мВ. В эксперименте с другой клеткой при частоте следования 2 и 5 импульсов в секунду величина деполяризации мембраны глиальной клетки составила, соответственно, 17 и 39 мВ ([1], с. 87).
|
![]()
 20 мВ
20 мВ
20 с
3. Деполяризация глиальных клеток, расположенных на расстоянии от нейрона, может возникнуть за счет распространения ионов К+ по щелевым контактам из ближних клеток в удаленные по концентрационному градиенту, что влечет выравнивание мембранного потенциала Ем в группе, состоящей из нескольких рядов сателлитов. Отсюда следует, что эта область в окружении нейрона, занятая деполяризованными глиальными клетками, может быть много больше размеров нейрона. Вывод подтверждается электрофизиологическим исследованием распространения медленного отрицательного потенциала (МОП), обусловленного деполяризацией глиальных мембран.
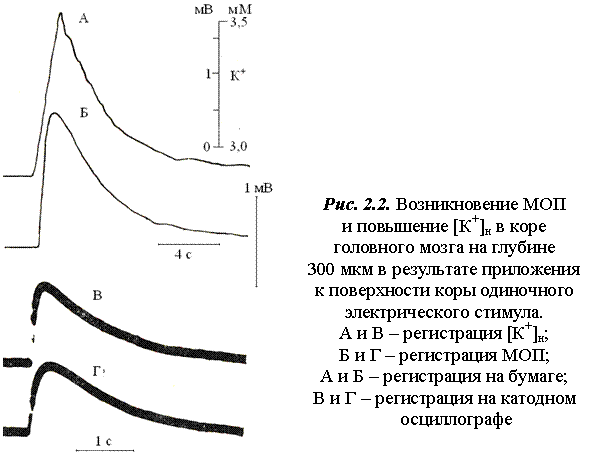
МОП регистрируется на поверхности и в глубине коры головного мозга одновременно с деполяризацией глиальных клеток (рис. 2.2). Реакция коры на прямое воздействие содержит два компонента: первый – быстрый, дендритный, возникает при малых значениях величины воздействующего импульса порядка 5-10 В и длительности 50-
100 мкс; второй, следующий за ним – МОП возникает при величинах стимула от 20 В и более.
Как принято в электрофизиологии возрастание величины МОП и внеклеточной концентрации ионов калия [К+]н, показано вверх от оси абсцисс (рис. 2.2 и далее).
О принадлежности МОП к процессам нейро-глиального взаимодействия на различных расстояниях от поверхности коры свидетельствуют результаты экспериментов.
На рис. 2.2 показано возникновение МОП в коре головного мозга в опыте с отводящими микроэлектродом, заполненным NaCl (регистрация МОП) и К+- селективным микроэлектродом, погруженными
в межклеточную жидкость на глубину 300 мкм. Развитие МОП
(рис. 2.2) коррелирует с повышением внеклеточной концентрации ионов калия [К+]н [20].
 |
2.2 Распределение медленного отрицательного потенциала
в коре головного мозга
МОП возникает в радиусе до 3, 5 мм от места нанесения стимула как при воздействии на поверхность коры, так и при воздействии на слои, лежащие на глубине от поверхности.
Прямые ответы коры вызывались в супрасильвиевой извилине кошки при глубоком нембуталовом наркозе (80-100 мг/кг живого веса). Раздражение – одиночный прямоугольный импульс длительностью 0,05 мс с амплитудой 50 В. Частота повторения воздействия – не более 1 имп/мин. Раздражающий двухполюсный электрод изготовлен из серебряной проволоки диаметром 0,1 мм; межполюсное расстояние – 0,4 мм. Отведение производилось стеклянным микроэлектродом, заполненным 2,5 М раствором NaCl. Использовался усилитель постоянного тока.
На рис. 2.3, 2.4А и 2.6А кривые распределения потенциала нормализованы: значение А величины МОП в данной точке поделено на ее максимальное значение А0. На рис. 2.4Б и 2.6Б приведены осциллограммы ответов коры.
В опыте (рис. 2.3) исследовалось распределение МОП на поверхности коры при раздражении электродом, установленным на супрасильвиевой извилине. Начальное расстоянии между раздражающим и отводящим электродами составляло 400 мкм. Величина МОП линейно снижалась по мере увеличения расстояния между ними. Полное исчезновение МОП произошло на расстоянии L=3100 мкм [21].
В серии из 17 экспериментов с послойным отведением медленного потенциала воздействие также производилось на поверхность супрасильвиевой извилины. Опыты различались только по расстоянию L между воздействующим электродом и проекцией отводящего электрода на поверхности коры, составлявшем от 0.5 до 1,6 мм.
На рис. 2.4 представлены результаты эксперимента, в котором расстояние между электродами равнялось 1 мм.
Как видно на рис. 2.4, с удалением отводящего электрода от поверхности коры амплитуда МОП немонотонно снижалась и на глубине hинв= 900 мкм снизилась до нуля. При дальнейшем погружении отводящего электрода до глубины ho= 3100 мкм регистрировался инвертированный (медленный положительный) потенциал. В области 600-800 мкм возникло "плато". В некоторых случаях плато могут быть плоскими (см область 100-160 мкм) или носить характер "горба" (область 260-360 мкм). В каждом эксперименте величина "горба" обычно не превышает 8-10% от максимального значения МОП - величины А0 . Ширина "плато" варьирует от 70 до 250 мкм; количество и расстояние от поверхности также изменчивы. Как правило, регистрируются одно или два "плато"; глубина их залегания – в районе первого слоя коры и перед точкой инверсии [22]*.
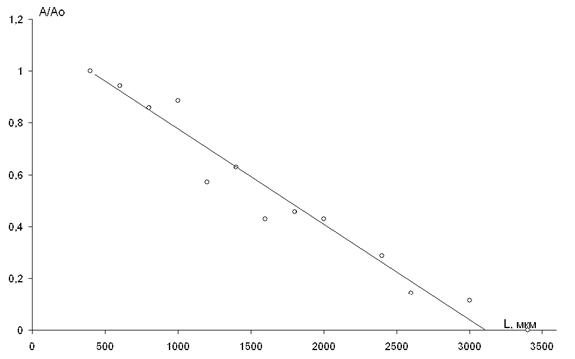
Рис. 2.3. Нормированная кривая распределения МОП на поверхности
супрасильвиевой извилины головного мозга наркотизированной
кошки при ее биполярном поверхностном раздражении.
Развертка – 1,5 с; калибровка – 1 мВ
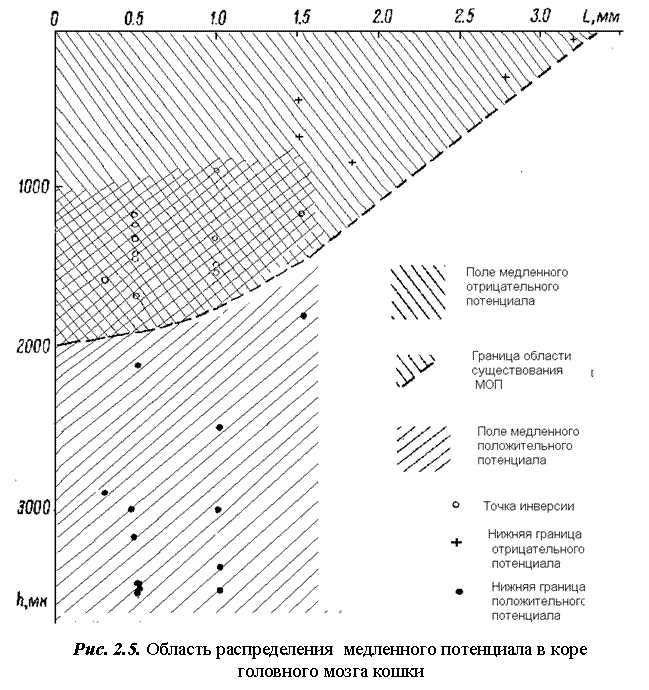
Обобщенные результаты 17 экспериментов приведены на рис. 2.5, на котором кружками обозначена глубина инверсии; крестиками – глубина исчезновения МОП; черными точками – глубина исчезновения инвертированного медленного потенциала.
В 12 экспериментах при расстоянии между раздражающим электродом и проекцией отводящего электрода на поверхности коры
L≤ 1,5 мм наблюдалась инверсия МОП. Из трех экспериментов, про-веденных при расстоянии L=1,5 мм, инверсия МОП возникла в одном
__________________________
*Необходимо учитывать, что толщина коры кошки много меньше толщины коры человека
случае на глубине 1150 мкм. В двух других полное исчезновение МОП произошло на глубине 480 и 720 мкм. При межэлектродных расстояниях свыше 1,5 мм (всего 3 эксперимента) инверсия МОП не возникала [22].
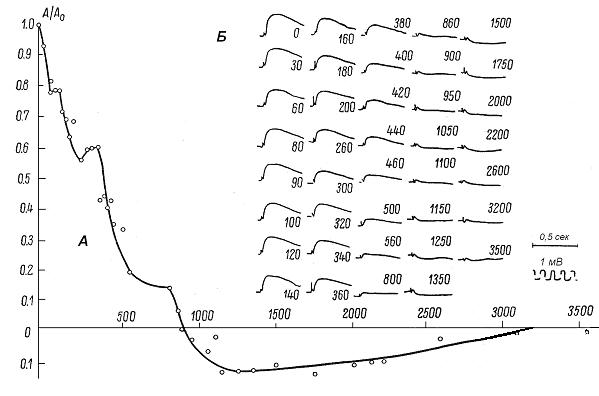
Рис. 2.4. Распределение медленного компонента прямого ответа
в глубине коры при расстоянии, равном 1 мм, между раздражающим
и отводящим электродами на поверхности.
Остальные условия те же, что на рис. 2.3
Результаты проведенной серии показали: все кривые распределения МОП в глубине коры делятся на два типа. К первому типу относятся кривые с инвертированным на некоторой глубине hинв. медленным отрицательным потенциалом. Большинство таких кривых получено при расстоянии между электродами до 1 мм (всего 12) и одна кривая при L=1,5 мм. Точка инверсии может находиться на глубине 900 ≤ hинв ≥1800 мкм.
Для кривых второго типа характерно отсутствие инверсии. Такие кривые получены при межэлектродных расстояниях L в области от 1,5 до 3,5 мм. МОП распространялся на глубину от 300 (L = 28 мм) до
900 мкм (L=1, 8 мм). Во всех случаях при дальнейшем погружении микроэлектрода медленный положительный потенциал не возникал.
 |
Амплитуда МОП может достигать 2,3 мВ. Амплитуда положительного потенциала колеблется в пределах 10-60 % от максимальной амплитуды МОП, но не бывает равной последней. Среднее ее значение для 10 проходов составляет 18,8 % от значения А0. Продолжительность МОП меняется от препарата к препарату в пределах от 0,35 до 3,5 с. Средняя продолжительность – 1,2 с. Определение длительности инвертированного потенциала часто затруднено из-за его малой величины. В некоторых случаях длительность МОП на поверхности коры и длительность положительного потенциала различны. На рис 2. 4Б, например, они разнятся примерно в два раза.
Итак, при воздействии на поверхность супрасильвиевой извилины головного мозга наркотизированной кошки МОП генерируется локально в области, расположенной под раздражающим электродом, и электротонически распространяется из места возникновения по поверхности и в глубину коры на расстояние в радиусе порядка 3-3,5 мм и 1,8 мм, соответственно.
При дальнейшей послойной регистрации возникает медленный положительный потенциал. Проекция поля этого потенциала на поверхности коры имеет меньшую, чем МОП, протяженность – порядка 1,5 мм. Эти показатели свидетельствуют о различных механизмах возникновения медленных отрицательного и положительного потенциалов, что допускает возможность взаимного перекрытия их полей. Можно предположить, что в области перекрытия в результате суперпозиции абсолютные значения их амплитудных показателей занижены, и сами поля медленного отрицательного и медленного положительного потенциалов, регистрируемые в глубинных слоях коры, простираются в перпендикулярном к поверхности коры направлении на расстояния большие, чем это следует из экспериментальных кривых послойного распределения.
По результатам экспериментов с раздражением поверхности коры, пространство, на которое распространяется МОП, можно представить как конусообразное тело вращения с основанием, лежащим на поверхности коры, и вершиной в нижних ее слоях на глубине 2-2,5мм. МОП возникает в точке приложения электрического стимула и распространяется электротонически в этом объеме по всем направлениям.
Поле положительного потенциала имеет форму цилиндра с осью, перпендикулярной к поверхности коры, высотой до 1,5-1,8 мм и диаметром основания до 2-х мм, расположенного в нижних слоях коры.
Таким образом, при послойном отведении истинное значение величины МОП, так же как и протяженность поля МОП, могут быть определены только на расстоянии L > 2-2,5 мм, на котором поле положительного потенциала отсутствует.
Приведенные на рис. 2.3-2.5 результаты не позволяют однозначно определить, генерируется ли МОП только в поверхностном слое коры или возникает в любой точке на расстоянии до 2-3 мм от поверхности. Для решения этого вопроса в серии экспериментов послойное раздражение и отведение производилось путем введения на заданную глубину супрасильвиевой извилины блока из трех покрытых эмалью металлических электродов, находившихся на расстоянии 0,6 мм друг от друга. Перед началом измерений в коре производился прокол на всю ее глубину и далее с выходом в подкорку. В результате перфорации МОП исчезал во всем объеме коры на расстоянии до 6 мм от перфорированного участка. Эксперимент начинался примерно через час после прокола, когда прямой ответ коры восстанавливался.
На рис. 2.6 представлен результат одного из экспериментов, из которого следует, что при воздействии на структуры коры, расположенные на разных уровнях относительно поверхности, и при регистрации ответа коры с этих же уровней, величина МОП последовательно снижается и исчезает полностью на глубине порядка 2,3 мм от поверхности коры. При дальнейшем погружении блока электродов в кору возникает медленный положительный потенциал, который регистрируется до глубины 4000 мкм. [23].
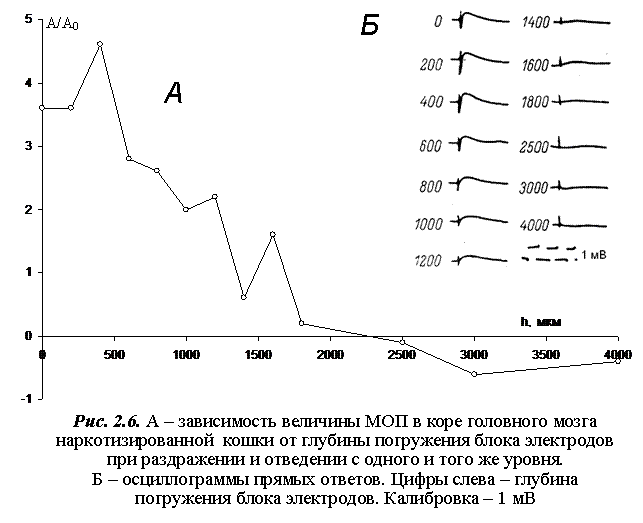 |
Итак, экспериментально показано, что МОП возникает в верхних слоях супрасильвиевой извилины головного мозга наркотизированной кошки в области, которая представляет собой конус с основанием, лежащим на поверхности коры и вершиной в глубине на расстоянии от 2-х до 3-х мм от поверхности коры. Максимальная активность процессов, связанных с генерацией МОП, локализована в области 300-400 мкм от поверхности коры, что соответствует ее II (наружному зернистому) слою.
Результаты послойного раздражения и отведения выявили ряд фактов, которые нельзя было обнаружить при поверхностном воздействии на кору. Эти факты играют важную роль при определении структур коры головного мозга, входящих в состав Процессора БКС.
Во-первых, обработка экспериментального материала показала, что максимум реакции на воздействие возникает не на самой поверхности коры, куда МОП распространяется электротонически, а на глубине порядка 400 мкм, т. е. активный процесс происходит в структурах коры головного мозга – во II слое коры головного мозга, в который поступает афферентная информация.
Во-вторых, установлено, что вопреки наличию описанных выше в п. 2.1 условий, необходимых для возникновения МОП (количество олигодендроцитов с глубиной не снижается, а возрастает), величина МОП непрерывно снижается по мере удаления раздражающего электрода от поверхности к нижним слоям коры.
В третьих, обнаружено, что при раздражении глубинных структур коры медленный положительный потенциал приобретает значительно большую протяженность, чем при раздражении поверхности коры.
Существенное отличие величины МОП в реакции, возникающей на глубине мкм, от величины МОП, регистрируемой на расстоянии 400 мкм от поверхности коры, свидетельствует о том, что при одинаковых параметрах воздействующего стимула деполяризация мембран глиальных клеток-сателлитов мелких звездчатых клеток IV (внутреннего зернистого) слоя чрезвычайно слаба или же отсутствует вовсе. Этот результат позволяет предположить, что до настоящего времени не выявлены все специфические особенности механизма возникновения МОП. Такой вывод подкрепляется спецификой исследований этого потенциала, которые проводились, в основном, на поверхности коры и в ее приповерхностных слоях. Отсутствие активного процесса деполяризации глиальных клеток в cлое IV явилось причиной высказанного выше в п. 1.3 заключения, согласно которому афферентный вход анализатора в сенсорной коре реализуется звездчатыми нервными клетками, расположенными во внешнем зернистом слое II.
Литература
1. Ройтбак и ее роль в нервной деятельности. «Наука», Ленинград, 1993.
2. Kornmuller A. E. Zum Wesen des EEG auf spezieller Untersuchungen: Congr. of neurologtical sci. (Brussels 1957) vol. III: EEG, clinical
neurophysiology and epilepsy. London etc., 1959. P. 243-247.
3. Watson W. E. Physiology of neuroglia//physiol. Rev. 1974, Vol. 54, p. 2
4. Brizee K. R., Vogt J., Kharetchko X. Postnatal changes in giia/neuron index with comparison of methods of cell enumeration in the white rat// Growth and maturation of the brain/Ed DP Purpura, J. M. Shadt, Amsterdam etc. 1964, Vol. 4, p. 136-149. (Progr. in brain res.)
5. Bennet E. L., Rosenzweig M. R., Krech D., Diamond M. C. Fur. ter evidance for a relation environmental complexity, learning abillity and number of glialcells//Physiologist. 1965, Vol. 8, p. 322.
6. Greer E. R., Diamond M. C., Tang J. M.W. Increase in thickness of cerebral cortex in responce to environmetal enrichment in Brattlebaro rats deficient in vasopressin// Exp. Neurol., 1981. Vol. 72, p. 366-379.
7. Diamond M. C., Scheibel A. B., Murphy G. M., Harvey T. On the brain of the Scientist Albert Enstein//Exp. Neurol. 1985. Vol. 88. P. 198-204/.
8. Eccls J. C. The anatomy of memory. Patio Alto, California, 1965.
p. 12.
9. Galambos R. A glia-neural theory of brain function//Procc. Nat. Acad. Sci. USA. 1961. Vol. 47. p. 129-136.
10. Galambos R. Introductory discussion of glial function//Progr. in brain res., 1965, Vol. 15. p. 267-277.
11. Purpura D. P. Brain and behavior/Ed. M. A. Brazier. Washington D. C. 1961. p. 190.
12. Swallow R. J. A neuron-glia cell model deduced from neurological and psichological considerations//Proc. 18-th annu. conf. eng. med. a biol. Philadelphia, 1975. P. 83.
13. (Pribram K. H.) Языки мозга. М., 19с.
14. Glees P. Neueroglia: Morphology and function. Illinois, 1955.
15. Аладжалова глиа-нейрональных поверхностей раздела в механизме краткосрочной памяти //1- съезд Всесоюзного физиологического общества им. . М. 1964. Т.2 С. 22-24.
16. Adey, W. R., Kado R. T., Mellwain J. T., Walter D. J. The role of neuronal еlements in regional cerebral impedance changes ina lerting,
orienting and discriminative responses//Exp. Neurol. 1966. Vol. 15. P. 490-510.
17. Adey. W. R. () Организация мозговых структур с точки зрения передачи и хранения информации//Cовременные проблемы электрофизиологии центральной нервной системы/Ред. , М., 1967. с. 324-340.
18. М,. И, Правдивцев центральной нервной системы. Академия. М., 2005, с. 54.
19. Kuffler S. W. Studies physiology of neuroglial cells //Proc. 24
Intern. Congr. Of physiol. Sci/ abdtractd. Washington,1968. Vol. 6. P.79-80
20. , , Очерашвили концентрации внеклеточного калия и медленный отрицательный потенциал в коре головного мозга. Нейрофизиология.
Т 12, № 5, Киев, 1980, с. 459-463.
21. , , О природе медленного отрицательного потенциала прямого ответа коры. Основные проблемы электрофизиологии головного мозга. «Наука», М., 1974 с.118-129.
22. Бобров поле в коре больших полушарий при медленном компоненте ее прямого ответа. Физиологический журнал СССР, 1973, Т 59, с. 378-383.
23. Бобров поле в коре больших полушарий при медленном компоненте ее прямого ответа. Физиологический журнал СССР, 1971, Т. 57. с. 656-663.
24. Бобров отрицательный потенциал в коре головного мозга при раздражении и отведении на одном уровне. Физиологический журнал СССР, 1975, Т. 61. с. 286-288.
Глава 3. ФУНКЦИИ НЕЙРОГЛИИ В МЕХАНИЗМЕ
СОЗНАНИЯ
3.1 Функция нейроглии в режиме переноса информации
на полевой уровень
В п. 1.4 показано, что условие непрерывного функционирования Процессора БКС обусловлено возможностью асинхронного поступления афферентной информации из рецепторных областей анализаторов в сенсорные области коры и далее в ассоциативные области. Сообщалось, что проблема перевода дискретной информации в аналоговую форму решается при участии деполяризованных глиальных клеток–сателлитов возбужденного нейрона, входящего в состав мозаики возбужденных нервных клеток, адекватно отображающей в корковой области анализатора образ внешней среды. При возбуждении нейрона деполяризованное состояние глиальной клетки-сателлита сохраняется на десятки секунд и даже на минуты (п. 2.1).
Механизм возникновения деполяризации клеток-сателлитов достаточно прост. При поступлении на синаптические входы нейрона афферентной информации нейрон возбуждается. При этом из межклеточного пространства в него поступают ионы натрия, а из клетки выходят ионы калия, которые аккумулируются в межклеточных щелях. Одновременно с ростом в межклеточном пространстве концентрации [К+]н возрастает градиент концентрации [К+]н относительно концентрации ионов [К+]вн в глиальной клетке. В результате она поглощает избыточный калий, что приводит к росту его внутриклеточной концентрации и, следовательно, к изменению значения величины равновесного мембранного потенциала Е0== –100 мВ до некоторого значения Ем (например, до Ем = –60 мВ), т. е. к деполяризации мембраны глиальной клетки. Таким образом, в ЦНС глиальная деполяризация клеток-сателлитов продолжительностью в десятки секунд является интегральным выражением нейронной активности в данном пункте коры. Это означает, что несмотря на прекращение поступления в Процессор информации о возбужденном состоянии нервной клетки, на полевом уровне сохраняется информация о пространственно-временной дислокации данного структурного элемента. Поступающая от рецептора новая афферентная информации, кодированная параметрами рецепторного потенциала, определит новое значение величины Ем у группы сателлитов, относящихся к данному нейрону. Это обеспечивает возможность обработки – прослеживания динамики, анализа и сравнения предшествовавшей афферентной информации с вновь поступившей из внешней среды.
3.2 Перенос информации с клеточного уровня на полевой
Рассмотрим еще раз подробнее механизм переноса афферентной информации, исходящей от рецепторных клеток какого-либо одного анализатора, с клеточного уровня на полевой. Такая информация поступает в сенсорную кору по нескольким каналам преимущественно от специфических сенсорных ядер таламуса, в которых она подвергается определенной обработке. Для упрощения задачи представим, что информация поступает только на мономодальные нервные клетки наружного зернистого слоя сенсорного поля коры головного мозга этого анализатора.
Афферентная информация, исходящая от i-й сенсорной клетки периферического конца анализатора, пройдя ряд переключательных нейронов подкорковых образований, поступает в i-ю колонку проекционной зоны коры (коркового конца анализатора по Павлову) и возбуждает в ней i-ю звездчатую клетку наружного зернистого (второго) слоя. Возбуждение i-го нейрона, входящего в состав афферентной мозаики, приводит к деполяризации группы окружающих его глиальных клеток-сателлитов. В результате диффузии К+ через щелевые контакты происходит деполяризация прилегающих к ним следующих удаленных от нейрона рядов клеток. В итоге, в некотором объеме вокруг возбужденного i-го нейрона мозаики возникает область, занятая группой клеток-сателлитов i-го нейрона с одинаковыми значениями величины отрицательного мембранного потенциала Ем, определяемого количеством поступивших в них ионов калия. Величина суммарного положительного заряда, поступившего в группу сателлитных клеток, и значение Ем адекватны параметрам афферентной информации на i-ом входе сенсорной зоны анализатора.
На полевом уровне БКС в Процессоре группа глиальных клеток-сателлитов i-го нейрона образует характеристическое поле, являющееся подсистемой характеристического поля сенсорной зоны коры и компонентом образа-афферента на уровне подсознания, адекватного информации на выходе i-ой рецепторной клетки анализатора.
Аналогичные группы глиальных клеток-сателлитов i-1 и i+1 нейронов, входящих в состав мозаики на уровне сенсорной зоны коры, образуют i-1 и i+1 подсистемы характеристического поля сенсорной зоны коры и, соответственно, i-1 и i+1компоненты образа-афферента на уровне подсознания.
Таким образом, всей афферентной информации, поступающей в каждый данный момент в проекционную (корковую) зону анализатора, на полевом уровне соответствует характеристическое поле деполяризованных глиальных клеток – сателлитов нейронов, участвующих в мозаике, которое содержит адекватную информацию, выраженную в аналоговой форме.
В случае поступления от рецептора на i-й нейрон только одного нервного импульса, характеристическое поле сателлитной группы этого нейрона сохраняется на секунды. При поступлении в тот же нейрон афферентной информации в виде серии импульсов, продолжительность существования характеристического поля, образуемого сателлитной группой этого нейрона, будет определяться суммой поступивших в нее положительных зарядов и скоростью выхода ионов калия после завершения серии. Результат суммации будет зависеть от частоты поступающих афферентных импульсов, их количества в серии и длительности самой серии, и может после интенсивной афферентации, постепенно затухая, сохраняться до нескольких минут, что на 4-6 порядков превышает длительность исходного нервного импульса. Понятно, что результаты суммации для каждого из компонентов мозаики будут непрерывно варьировать в зависимости от поступающей к каждому из них афферентной информации. Эти показатели совпадают с характеристиками кратковременной памяти.
Итак, на уровне характеристического поля глиальной клетки информация кодирована в виде количества ионов калия, определяющих значение ее мембранного потенциала Ем, адекватного афферентной информации, поступающей на нейрон от рецептора. Значение этого потенциала сохраняется у всех глиальных клеток, входящих в группу сателлитов. Площадь, занимаемая сателлитной группой, меньше площади, упомянутой в п. 1.1 нейронной колонки, являющейся представительством одного рецептора данного органа. Таким образом, каждый фрагмент мозаики нейронов на уровне Процессора строго детерминирован как по величине, так и по его дислокации.
На полевом уровне биокомпьютера сознания – Процессора – афферентная информация, поступающая в сенсорные зоны анализаторов, будет представлена характеристическим полем образа-афферен-та, адекватным распределению концентрации ионов калия в сателлитных группах всех возбужденных нейронов мозаики. На уровне физического вакуума она представлена фантомом образа-афферента – образом*. Продолжительность существования образа-афферента определяется временными параметрами процессов, проходящих во внешней среде; продолжительность существования фантома образа не ограничена.
Как следует из всего сказанного, временные характеристики образа-афферента коррелируют с временными характеристиками кратковременной памяти, а временные характеристики фантома образа коррелируют с временными характеристиками долговременной памяти, что позволяет судить о полевой природе феноменов кратковременной и долговременной памяти и их механизмах.
Подведем итог: Проецирование на входы сенсорных зон анализатора афферентной информации, адекватной образу внешней среды, приводит к возникновению афферентной мозаики возбужденных нейронов внешнего зернистого слоя и к последующей деполяризации глиальных клеток, входящих в группы их сателлитов. Деполяризация обусловлена поглощением этими клетками ионов калия, выходящего из возбужденных нейронов мозаики в межклеточные щели.
На уровне Процессора характеристическое поле деполяризованных глиальных клеток, входящих в состав сателлитных групп,
содержит образ-афферент, адекватный образу внешней среды.
На уровне физического вакуума образу внешней среды соответствует фантом образа в архивах долговременной памяти.
Из всего сказанного в п. 3.2 следует: участие глиальных клеток коры головного мозга в процессах обработки афферентной информации на полевом уровне заключается:
1. В переводе дискретной информации, поступающей в кору из подкорковых отделов головного мозга в аналоговую форму.
2. В процессах обработки афферентной информации, поступающей от специфических рецепторных систем, систем жизнеобеспечения и из физического вакуума.
3. В участии в механизмах кратковременной и долговременной памяти.
_________________________
* Такое искусственное разделение введено для упрощения усвоения материала. В действительности, характеристические поля материальных объектов также принадлежат физическому вакууму
Глава 4. ФУНКЦИОНИРОВАНИЕ БИОКОМПЬЮТЕРА
СОЗНАНИЯ
Головной мозг (ГМ), являясь главным отделом центральной нервной системы, обеспечивает регуляцию процессов во всех системах жизнеобеспечения организма в условиях окружающей среды. Одновременно ГМ является субстратом психической деятельности высших животных и человека, в частности его сознания. При бодрствовании функционирование ГМ происходит в условиях постоянного поступления афферентной информации в нижние отделы, в сенсорные и ассоциативные области коры. Афферентная информация исходит от раз-личных сенсорных систем (рецепторов), отражающих состояние окру-жающей среды и самого организма, а также из физического вакуума.
К сенсорной функции головного мозга относится сохранение афферентной информации, поступающей из окружающей среды, оценка ее биологической значимости, анализ и отбор на основе имеющегося опыта (памяти). Сенсорная функция реализуется при участии анализаторов, каждый из которых состоит из периферической части (системы рецепторов – источников афферентной информации), каналов ее передачи и проекционной зоны коры (корковых отделов анализаторов). Проекционные зоны коры расположены в различных областях неокортекса, в которые афферентная информация поступает после многократных переключений и придания ей нового качества (модальности), например эмоциональной окраски, проведенного в нижних отделах головного мозга.
По выполняемым функциям и морфологическим признакам в каждом анализаторе различают три связанные между собой проекционные зоны: первичную (центральное проекционное поле), состоящую из мономодальных нейронов; вторичную, расположенную на периферии относительно центрального поля; и ассоциативную, входящую в состав ассоциативной области коры.
На полимодальные нервные клетки ассоциативной области коры (области перекрытия анализаторов, например слухового и зрительного) информация поступает из вторичных проекционных зон анализаторов, а также из нижних отделов головного мозга, главным образом, от ассоциативных ядер таламуса. При этом в ассоциативных корковых проекционных зонах каждого из анализаторов возникают мозаики возбужденных нейронов, которые на полевом уровне (в Процессоре) при посредстве характеристических полей деполяризованных глиаль-
ных клеток-сателлитов представляют адекватные образы-афференты.
Каждый из таких образов является подсистемой характеристического поля ассоциативной области коры, несущего информацию о спиновой системе сложного образа-афферента, поступившего из периферических частей двух анализаторов. Это позволяет на уровне Процессора при активном участии Аппарата афферентного синтеза производить сравнительный анализ, оценку значимости, отбор, коррекцию и другие операции над поступающей в анализаторы афферентной информацией.
Кроме информации о состоянии внешнего мира, проецируемой в сенсорную кору от специализированных рецепторов, на полевой уровень БКС поступает информация из физического вакуума, в том числе из архивов долговременной памяти. По таламо-кортикальным путям поступает также афферентная информация от систем, обеспечивающих регуляцию процессов жизнедеятельности самого организма и его безопасность. На уровне Процессора характеристические поля, несущие информацию о спиновых системах областей, содержащих эту информацию, являются подсистемами спиновой системы коры головного мозга.
4.1 Возникновение образов, сложных образов и идей. Перенос
их на полевой уровень и в архивы долговременной памяти
Материальным субстратом Процессора являются все те области неокортекса, в которые поступает афферентная информация из нижних отделов головного мозга и с уровня физического вакуума и где происходит сравнительный анализ, оценка и отбор образов, сложных образов и идей, возникших на их основе. К ним относятся как сенсорные области, в которые проецируются все внешние и внутренние раздражители, так и ассоциативные области коры.
На уровне Процессора клеточные структуры этих областей представлены характеристическими полями, несущими информацию о структуре их спиновой системе. Спиновые системы характеристических полей в каждый данный момент определяются их информационным наполнением. На уровне физического вакуума эти структуры представлены торсионными полями, несущими информацию о характеристических полях в областях сенсорной и ассоциативной коры.
Если принять, что информационное наполнение какой-либо сенсорной области коры определяется только афферентной информацией, поступающей от рецепторов, отражающих некий образ внешней среды, то на уровне Процессора мозаике возбужденных нейронов-афферентов этого образа, адекватно соответствует характеристическое поле образа-афферента внешней среды; на уровне физического вакуума – торсионное поле-фантом образа-афферента
Долговременная память представлена на уровне физического вакуума торсионными архивами образов. Все образы-афференты, как сенсорных областей анализаторов, так и ассоциативных областей, возникшие на уровне Процессора, сохраняются на уровне физического вакуума в архивах образов долговременной памяти в качестве торсионных фантомов характеристических полей деполяризованных глиальных клеток, содержащих информацию о мозаике возбужденных нейронов. Афферентная информация, непрерывно поступающая в сенсорные области коры, доставляет образы в долговременную память в виде торсионных фантомов простых образов. Аналогично, в каждый данный момент архивы сложных образов долговременной памяти пополняются торсионными фантомами характеристических полей деполяризованных глиальных клеток, содержащих информацию о мозаике возбужденных нейронов в ассоциативных областях неокортекса. В процессе анализа и оценки значимости образов и сложных образов, происходящих при участии накопленного опыта (информации, поступающей из архивов долговременной памяти), возникают "идеи", торсионные фантомы которых также сохраняются на уровне физического вакуума в "архивах идей".

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 |
