
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Лісорослинні умови також впливають на розподіл сумарної активності 137Cs в біогеоценозі. При зниженні трофності грунту концентрації радіонуклідів у лісовій рослинності зростають. В умовах складних суборів загальний запас радіонуклідів у деревостані втроє менший, ніж у суборових і в 4-5 разів менший, ніж у борових умовах місцезростання. Аналогічна ситуація характерна і для гігротопів. Найвища акумуляція радіонуклідів у деревостані у сирих умовах (більше 30 % загального запасу біогеоценозу). У вологих вона менша вдвоє, а в свіжих — в 4 рази.
Особливості міграції радіонуклідів в екосистемах лісових боліт
Особливе місце в міграції радіоізотопів у довкіллі посідають лісові болота. Як відомо, ці екосистеми міцно утримують значну сумарну активність радіонуклідів, є біогеохімічними бар'єрами на шляху їх міграції у грунтово-рослинному покриві взаємопов'язаних ландшафтів. За характером мінерального живлення та рослинним покривом лісові болота об'єднують у три типи:
- верхові (оліготрофні);
- перехідні (мезотрофні);
- низинні (евтрофні).
Міграція радіонуклідів у лісових болотах відбувається переважно у системі: "субстрат (торф) - ґрунтова вода - рослинний покрив". У наведених вище типах боліт міграція радіонуклідів істотно відрізняється. У верхових та перехідних болотах вона приблизно однакова. Це зумовлено наявністю майже суцільного сфагнового покриву, переважанням сосново - або березово-чагарниково-сфагнових та трав'яно-сфагнових ценозів, низькими значеннями кислотності (pH=3,0-4,00) і мінералізації ґрунтової води, низьким ступенем розкладу торфів (20-35 %) та їх об'ємною вагою (0,08 г/см3). Значна різниця спостерігається між названими двома типами боліт та низинними болотами. Для останніх характерна висока мінералізація водних розчинів, їх слабокисла або нейтральна реакція (рН=5,0-7,0), інтенсивніший водообмін, часто - проточний режим зволоження. Рослинний покрив їх складається переважно з вільхово-трав'яних, рідше -трав'яно-гіпнових угрупувань. Для їх торфів характерний високий ступінь розкладу (до 80 %) та більша (до 0,2 г/см3) об'ємна вага, ніж у торфів боліт попередньої групи.
Специфічну роль у міграції радіонуклідів у болотних біогеоценозах відіграє вода, яку за локалізацією у певному елементі болотної екосистеми поділяють на чотири типи:
- імпрегновану, що утримується рослинним покривом;
- болотну, що формує відкрите водне дзеркало;
- ґрунтову (торфову, порову), що утримується торфами;
- дренажну (стокову), що міститься у меліоративній мережі.
-
За вмістом 137Cs наведені типи можна розмістити у такий ряд: порові > імпрегновані > болотні > дренажні. Однак, як правило, вміст радіонуклідів у всіх типах вод лісових боліт у 100-1000 разів менший, ніж у торфах. Це пояснюється значною катіонною місткістю торфів щодо радіонуклідів, зокрема, 137Cs. Однак у той же час для екосистем боліт характерний відносно високий вміст мобільних форм радіонуклідів у грунтах (торфах). Тому основні довгоживучі радіонукліди у грунтово-рослинному покриві лісоболотних екосистем за постійного перезволоження екотопів мають більшу міграційну здатність порівняно з лісовими біогеоценозами суходолів. Це явище також посилюється незначним вмістом у торфах мінеральної складової грунту (мулистої та глинистої фракцій), на якій сорбуються радіонукліди (зокрема 137Cs) у мінеральних грунтах, нестачею основних макроелементів - азоту, калію, фосфору, доступних для кореневого живлення рослин.
Вертикальна міграція радіонуклідів у торфах відбувається у 5-7 разів швидше, ніж у піщаних грунтах суходольних лісів. Для торфів характерним є специфічний вертикальний розподіл радіонуклідів. Зокрема, більша частина валового запасу l37Cs міститься у верхньому 20-см шарі, однак максимум вмісту радіонукліда переміщується з шару 0-1 см (1986 р.) у шар 4-6 (8) см (1996р.). У той же час для 90Sr характерний більш-менш рівномірний вертикальний розподіл у торфі.
У лісоболотних екосистемах значну роль у транспортуванні основних елементів живлення і радіонуклідів до рослин відіграє мікориза. Значення її в цих процесах зростає від низинних боліт до верхових. Серед рослин усіх континентальних біогеоценозів Європи саме судинні рослини лісових боліт (особливо мезотрофних та оліготрофних) найбільш інтенсивно накопичують радіонукліди. Наприклад, у 1997 р. на мезотрофній ділянці болотного масиву "Бучмани" (Житомирська область) у ценозі сосняка чорнично-пухівково-сфагнового при щільності забруднення грунту '37Cs 39 кБк/м2 (близько 1,0 Кі/км2) питома активність радіонукліда в надземній фітомасі видів трав'яно-чагарничкового ярусу в середньому становила:
рінхоспори білої - 23000 Бк/кг,
журавлини - 13600 Бк/кг,
чорниці - 11400 Бк/кг,
буяхів - 9300 Бк/кг,
соки попелястої -7500 Бк/кг,
підбілу білолистого - 4800 Бк/кг.
Рослини евтрофних лісових боліт менш інтенсивно накопичують радіонукліди, ніж оліготрофних та мезотрофних.
З радіоекологічного погляду істотною особливістю боліт є те, що інтенсивний малий (біологічний) кругообіг радіонуклідів тут відбувається швидше порівняно з великим (геологічним). Інтенсивне накопичення радіонуклідів у рослинному покриві, його значна біомаса зумовили нерівномірність розподілу валового запасу радіонуклідів між грунтом (торфом) та рослинним покривом. Нині саме в рослинному покриві лісоболотних екосистем зосереджена більша частина сумарної активності радіонуклідів, зокрема, 137Cs.
Міграція радіонуклідів в екосистемах верхових та перехідних боліт має спільні риси, пов'язані з наявністю щільного (з покриттям до 90-98 %) та потужного (10-30 см завтовшки) сфагнового покриву. При аеральному забрудненні оліготрофної лісоболотної екосистеми 137Cs, радіоактивні частинки лише незначною мірою були затримані кронами сосни (внаслідок невеликої їх зімкнутості - 0,4-0,5) та трав'яно-чагарничковим ярусом. Більша ж частина активності 137Cs сорбувалася безпосередньо сфагновим покривом. А через 2-3 місяці на його поверхню ще додатково надійшла значна частка радіонукліда з верхніх ярусів рослинності.
Протягом першого вегетаційного періоду після аварії близько 90-95 % сумарної активності міцно утримувалося у шарі живих сфагнів. Після закінчення вегетаційного періоду, коли фізіологічна активність сфагнів різко зменшилася, частина активності радіонукліда мігрувала донизу - у шар мертвих сфагнів, очосу, у верхній шар власне торфу. Згаданий низхідний потік радіонуклідів (за осіннього гідрологічного піку) був досить інтенсивним. До початку вегетації наступного року спостерігається ще один гідрологічний пік весняний, пов'язаний із сніготаненням. У період цих гідрологічних піків обводнення оліготрофних та мезотрофних боліт істотно зростає, а фізіологічна активність рослинного покриву, в тому числі й сфагнів, дуже незначна внаслідок низьких температур повітря та субстрату. Саме в ці періоди l37Cs максимально вилуговується із рослинності, особливо сфагнів. Це зумовлює його вищу концентрацію у всіх видах води боліт порівняно з вегетаційним періодом.
Початок вегетації кардинально змінює ситуацію. Запас 137Cs, що містився в імпрегнованій та частково у поровій воді, а також у відмерлих частинках сфагнів, у зв'язку з фізіологічними особливостями цих мохів надходить до їх верхівкових, живих частин. Висхідний потік зумовлений не пасивним капілярним підняттям води до верхніх частин мохів, а активною іонообмінною дією клітинних оболонок живих частин сфагнів. У цей час концентрація 137Cs у воді боліт різко зменшується і утримується на низькому рівні (1-3 Бк/л) практично протягом усієї вегетації. Вилуговування 137Cs у нижні горизонти сфагнів та торф протягом гідрологічних піків у холодну пору року, повернення значної активності радіонукліда у верхню, живу частину сфагнового покриву протягом вегетації відбувається щорічно. Види ж трав'яно-чагарничкового ярусу верхових боліт поглинають радіонуклід з імпрегнованої та порової води, очосу, торфу. Разом з деревним наметом вони виносять радіонуклід з більш глибоких шарів торфу, ніж сфагнові мохи.
На відміну від оліготрофних та мезотрофиих боліт, для яких характерними є відсутність лісової підстилки (вона захороняється щорічно внаслідок наростання сфагнів) та повільний (6-8 років) розклад відпаду, в евтрофних болотах істотно зростає швидкість мінералізації рослинних решток. У болотах цього типу лісова підстилка щорічно формується і розкладається протягом 1-2 років. Крім того, на низинних болотах посилюється надходження радіонуклідів з водним потоком, адже часто ці болота розміщуються у заплавах та присхилових ділянках. При цьому радіонукліди в екосистему можуть надходити не лише аеральним шляхом, але й з поверхневими та поровими водами. Основні закономірності міграції 137Cs в екосистемі низинного болота добре ілюструються на прикладі заплавного вільшняка осокового (рис.).
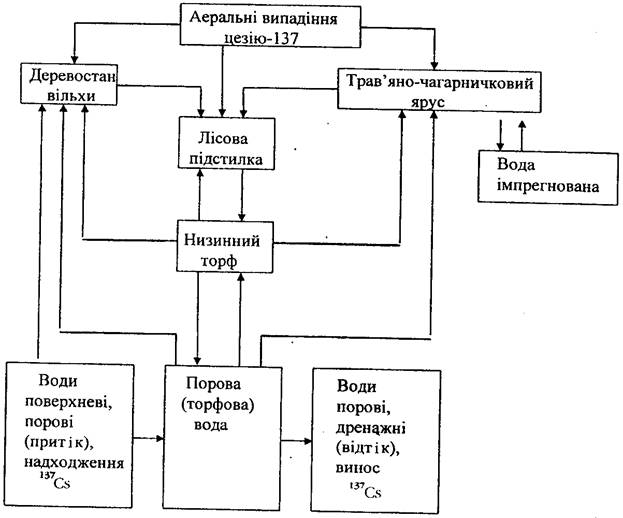
Рис. 7.7. Схема міграції 137Cs в евтрофній болотній екосистемі вільхового лісу осокового.
Для цього ценозу типовим є: деревостан вільхи із зімкнутістю 0,7, трав'яно-чагарничковий ярус з проективним покриттям 60 %, відсутність мохового покриву. Радіонукліди, зокрема l37Cs, надходять у цю екосистему переважно аеральним шляхом і 50-60 %їх активності затримується кронами дерев. Через 2-3 місяці, внаслідок самоочищення крон, значна частка активності 137Cs переміщується на поверхню трав'яно-чагарничкового ярусу та торфу. Після закінчення вегетації, листопаду дерев та відпаду надземної фітомаси трав більша частка сумарної активності радіонукліда евтрофної лісоболотної екосистеми була вже у лісовій підстилці. За швидкої мінералізації підстилки через рік значна кількість радіонукліда переміщується до верхнього, коренезаселеного шару грунту. З останнього кореневим шляхом l37Cs інтенсивно надходить до рослинного покриву деревного та трав'яно-чагарничкового ярусів. Для низинних боліт типова інтенсивна міграція радіонуклідів у системі "грунт-рослина-грунт", цикл якої повторюється щорічно.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 |
