
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
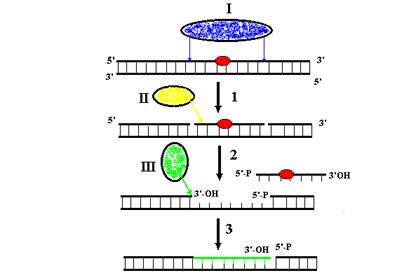
Рис. 1.4. Репаративный синтез ДНК в процессе нуклеотидной эксцизионной репарации.
1 – инцизия поврежденного участка нити ДНК эксцизионными эндонуклеазами, 2 – расплетание сегмента нити ДНК между ОР ДНК-лигазой и освобождение этого сегмента из дуплекса, 3 – репаративный ресинтез однонитевой бреши ДНК.
I – эксцизионная эндонуклеаза, II – ДНК-геликаза, III – ДНК-полимераза
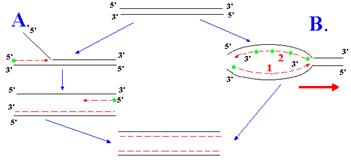
Рис. 1.5. Непрерывная (А) и полунепрерывная (В) репликация ДНК.
Родительские нити ДНК изображены сплошными черными линиями, вновь синтезированные нити – прерывистыми красными линиями, а праймеры РНК – зелеными кружками. Тонкие красные стрелки указывают направление роста вновь синтезируемых нитей ДНК, а толстая красная стрелка – направление движения репликативной вилки. На рис. В красными цифрами обозначены ведущая нить (1) и отстающая нить (2)
При полунепрерывной репликации одна из дочерних нитей элонгируется непрерывно из одного праймера, использованного при инициации (рис. 1.6, В). Эта нить ДНК называется ведущей. Полимеризация непрерывной нити идет в том же направлении, в котором перемещается по дуплексу родительской ДНК репликативная вилка. Нить дочерней ДНК, комплементарная ведущей, называется отстающей. Синтез отстающей нити происходит одновременно с синтезом ведущей нити, но в направлении, противоположном движению репликативной нити. Такой синтез не может быть непрерывным, т. к. ДНК-полимеразы перемещаются по матричной нити ДНК только в одном направлении 3’®5’. Отстающая нить создается из дискретных коротких цепей ДНК, синтезируемых на матрице для этой нити в результате перемещения ДНК-полимеразы в стандартном направлении. Эти короткие цепи называются фрагментами Оказаки. Каждый из таких фрагментов должен инициироваться заново из праймеров РНК, синтезируемых праймазой. При полунепрерывной репликации для завершения синтеза отстающей нити требуется участие «репаративной» системы, которая удаляет праймеры РНК, заполняет образующиеся бреши и соединяет друг с другом короткие отрезки вновь синтезированной ДНК. В процессе полунепрерыной репликации участвуют сложные многокомпонентные машины репликации, называемые реплисомами. Сборку и работу этих машин у прокариотов и эукаритов мы рассмотрим с гл. 2-3. Однако вначале необходимо остановиться на свойствах основных типов ДНК, полимераз, участвующих в репликации и репарации.
1.2. Бактериальные ДНК-полимеразы
Характеристики главных типов бактериальных ДНК-полимераз наиболее детально изучены на примере E. coli (табл. 1.2). Всего у кишечной палочки существуют 5 разных ДНК-полимераз. Мы рассмотрим вначале только три из них, отложив характеристику двух «склонных к ошибкам» ДНК-полимераз до гл. 00.
Таблица 1.2
ДНК-полимеразы E. coli
ДНК-полимеразы | Ген | Положение на карте, мин | Продукт | Число остатков | Мол. масса кД | Функция |
ДНК-полимераза I (A) | polA | 87,2 | PolI | 928 | 103 | Репарация, репликация |
ДНК-полимераза II (B) | polB | 1,4 | PolII | 783 | 90 | Репарация, мутагенез |
Главная репликативная ДНК-полимераза III (C) | dnaE | 4,4 | a, PolIII | 1160 | 132 | Каталитическая субъединица |
dnaN | 83,6 | b, PolIII | 366 | 41 | Скользящий зажим | |
dnaQ | 5,1 | e, PolIII | 243 | 27 | (3’®5’)-экзонуклеазная субъединица | |
dnaX | 10,6 | t, PolIII g, PolIII | 643 | 71 | Структурная субъединица | |
431 | 47 | Комплекс g-комплекса погрузчика зажима | ||||
holA | 14,4 | d, PolIII | 343 | 39 | “ | |
holB | 24,9 | d’, PolIII | 334 | 37 | “ | |
holC | 96,6 | c, PolIII | 147 | 17 | “ | |
holD | 99,3 | y, PolIII | 137 | 15 | “ | |
holE | 41,5 | q, PolIII | 76 | 8 | Компонент минимального фермента |
1.2.1. ДНК-полимераза I E.coli
Из всех ДНК-полимераз E. coli наиболее высокий внутриклеточный уровень (~ 400 молекул на клетку) имеет ДНК-полимераза I, кодируемая существенным геном polA. Она состоит из одной субъединицы длиной 928 остатков, которую можно разделить на три домена (рис. 1.6). N-концевую треть молекулы занимает (5’®3’)-экзонуклеазный домен, в центре расположен самый короткий (3’®5’)-экзонуклеазный домен, а самый большой полимеразный домен находится на С-конце. При ограниченном протеолизе ДНК-полимеразы I расщепление на границе между первыми доменами дает малый N-концевой 5’-нуклеазный фрагмент и большой С-концевой фрагмент, который называется кленовским фрагментом. Последний сохраняет 3’-нуклеазную и полимеразную активность и часто используется для синтеза ДНК in vitro.
(3’®5’)-экзонуклеазный домен, ответственный за корректорскую активность ДНК-полимеразы I, не является обязательным и отсутствует у бактерий из родов Thermus и Rickettsia. Остальные два домена жизненно важны для клеток и обеспечивают участие ДНК-полимеразы I в процессах репарации и удаления рибонуклеотидной части фрагментов Оказаки в отстающей нити. Поэтому делеция гена polA летальна для E. coli, а нелетальные мутации понижают в 10 раз скорость соединения фрагментов Оказаки и повыщаютповещают чувствительность клеток к агентам, повреждающим ДНК.
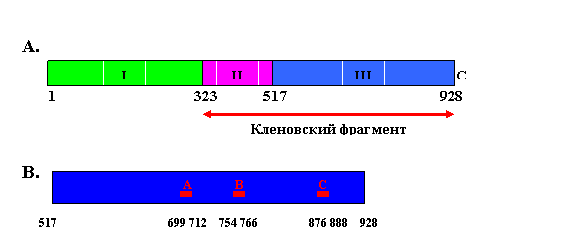

Рис. 1.6. Доменная с
труктура ДНК-полимеразы I E.coli.
A – взаимное расположение доменов. I – 5’-нуклеазный домен, II – (3’®5’)-экзонуклеазный домен, III – ДНК-полимеразный домен. Стрелка соответствует кленовскому фрагменту ДНК-полимеразы I.
B – мотивы полимеразного домена, консервативные среди бактериальных ДНК-полимераз I.
Эти мотивы имеют следующие консенсусные последовательности: 699hhhhDhhxEhx712 (A), 754RpxxKxxxhGhhY766 (B) и 876hhxhHDЕhxxЕ888 (С) - , где h – гидрофобный остаток, х – произвольный остаток, р – преимущественно R (нумерация остатков ДНК-полимеразы I E. coli)
Полимеразные домены различных ДНК-полимераз I и ДНК-полимеразы фага Т7 очень похожи друг на друга и содержат 6 консервативных участков, среди которых три аналогичны консервативным мотивам А, В и С главного семейства В эукариотических ДНК-полимераз и играют наиболее важную роль во время синтеза ДНК. Мотивы А и С ДНК-полимеразы I E. coli содержат кислые остатки асп705 и асп882 соответственно, связывающие катионы Mg+ в каталитическом активном центре (см. рис. 1.1В), а мотив В включает инвариантный положительно заряженный остаток (арг758) и ароматический остаток (тир766). Эти три мотива являются элементами полости, с которой связывается включающийся дНТФ в активном полимеразном центре.
-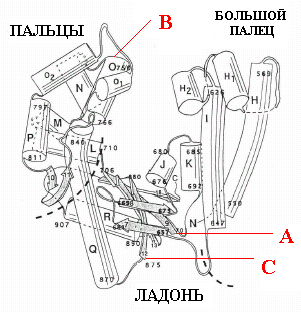
Рис. 1.7. Модель трехмерной структуры полимеразного домена ДНК-полимеразы I E.coli.
Указаны положения, в которых начинаются консервативные мотивы А, В и С
Рентгеноструктурный анализ кленовского фрагмента ДНК-полимеразы I E. coli впервые обнаружил специфическую архитектуру полимеразного домена, который по форме напоминает кисть правой руки (рис. 1.7) и состоит из пространственных доменов «ладони» (palm), «большого пальца» (thumb) и «пальцев» (fingers). Аналогичную трехмерную организацию имеют и все изученные экспериментально полимеразные домены других ДНК-полимераз и обратной транскриптазы вируса иммунодефицита человека. Можно предположить, что так же организованы и полимеразные домены большинства ДНК-полимераз. Наиболее консервативна топология домена ладони, состоящего из b-слоя, фланкированного двумя a-спиралями. В домене ладони расположены консервативные мотивы А и С. Домены большого пальца и пальцев у ДНК-полимеразы I E. coli образованы a-спиралями, причем спираль О домена пальцев совпадает с мотивом В. У других ДНК-полимераз организация этих доменов гораздо менее консервативна. Хотя домен больших пальцев остается преимущественно a-спиральным, но тонкие детали его топологии неодинаковы. Еще более велики различия доменов пальцев. В целом, для архитектуры полимеразного домена характерно существование внутренней полости для связывания матрицы-затравки ДНК и входящего дНТФ, основанием которой является домен ладони.
На примере бактериальной ДНК-полимеразы I мы рассмотрим последовательные изменения конформации этой полости во время полимеразного каталитического цикла. Эти рентгеноструктурные данные были получены для нескольких свободных бактериальных полимераз и их комплексов с ДНК в присутствии и в отсутствие дНТФ. Вероятно, такие же изменения происходят во время работы других ДНК-полимераз. «Рабочий цикл» состоит из 4 последовательных стадий: 1) связывания ДНК-полимеразы с матрицей-затравкой ДНК; 2) связывания с комплексом полимераза-ДНК подходящего дНТФ, комплементарного первому остатку матрицы; 3) нуклеофильной атаки, приводящей к образованию фосфодиэфирной связи; 4) освобождения пирофосфата. Первые две стадии протекают очень быстро, и общую скорость полимеризации лимитирует стадия 3 или предшествующее ей изменение конформации.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 |
