Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Сравнивая показатели интенсивности течения ПОЛ у коров с кистами и гипофункцией яичников следует отметить, что у последних имеет место более выраженная функциональная недостаточность как ферментативного, так и неферментативного звеньев антиоксидантной защиты. При этом концентрация продукта перекисного окисления – МДА – у них оказалась выше на 20,8%, а ГПО – ниже на 16,7%, ГР – на 5,6%, СОД – на 17,9%, каталазы – на 7,4%, витамина Е – на 74,3%. Можно допустить, что снижение мощности антиоксидантной защиты у данных животных связано с дефицитом селена в их организме. У клинически здоровых коров его концентрация в крови составила 16,0±0,73 мкг%, у коров с кистозным поражением яичников – 14,6±0,68 мкг%, а с гипофункцией яичников – 12,7±0,32 мкг%.
Таблица 9
Некоторые показатели состояния системы ПОЛ-АОЗ у циклирующих и
бесплодных коров с патологией яичников
Показатель | Коровы здоровые циклирующие | Коровы бесплодные с | |
кисты яичников | гипофункция яичников | ||
МДА, мкМ/л | 1,00±0,05 | 1, 30±0,09 | 1,57±0,06 |
ГПО, мМ G-SH/л´мин | 9,4±0,32 | 14,0±0,55 | 12,0±0,64 |
ГР, мкМ G-SS-G/л´мин | 293,0±10,88 | 342,0±6,89 | 324,0±8,24 |
СОД, усл. ед./мг Hb | 0,72±0,03 | 1,12±0,04 | 0,95±0,05 |
Каталаза, мМ Н2О2/л´мин | 25,9±0,57 | 34,6±0,98 | 32,2±0,84 |
Витамин Е, мг% | 1,02±0,15 | 1,22±0,23 | 0,70±0,12 |
Каротин, мкг% | 476,0±37,1 | 577,0±57,1 | 326,5±58,9 |
Полученные при оценке интенсивности течения процессов перекисного окисления липидов у бесплодных коров с дисфункцией яичников данные позволяют также предположить, что в нарушении стероидогенеза у таких животных, характерном для данной патологии, могут лежать и процессы усиленного течения свободнорадикального окисления (Н. К. Зенков и др., 2001).
При оценке состояния системы оксида азота у коров с гипофункцией яичников также установлено (Г. Н. Близнецова, В. А. Сафонов), что им свойственен низкий уровень концентрации в крови как стабильных метаболитов NO· (20,6±2,21 мкМ/л против 52,9±2,46 мкМ/л), так и S-нитрозотиолов (2258,0±34,3 нМоль/мл против 3215,0±101,0 нМоль/мл). Разница в содержании NO· составила 2,57 раза (Р<0,001), а S-нитрозотиолов – 1,42 раза (Р<0,001).
Поскольку оксид азота принимает участие в механизмах антиоксидантной защиты (S. J. Klebanof, 1992; K. Dobashi et al., 1997; S. Ulker et al., 2003), то можно допустить, что недостаточная его генерация в организме животных (возможно в связи с ингибированием NO·-синтаз) также является одной из причин возрастания скорости свободнорадикального окисления.
Исходя из того, что некоторыми авторами установлена взаимосвязь образования в организме NO· с уровнем концентрации эстрогенов (A. S. Moreno, C. R. Franci, 2004) и прогестерона (T. Coughlan et al., 2005), можно предположить, что снижение продукции NO· у коров с патологией яичников связано с низким уровнем концентрации половых стероидов в их организме (рис. 1).
Не исключено, что наблюдаемое нарушение процесса фолликуло - и стероидогенеза у коров при дисфункции яичников связано, в свою очередь, с низким содержанием в их организме оксида азота, так как в работе P. T. Gond et al. (2008) показано, что данное соединение способно замедлять старение половых клеток (яйцеклеток), жизнедеятельность которых определяет рост, созревание фолликулов и функциональную активность яичников. То есть, низкий уровень оксида азота ведет к ускорению старения яйцеклетки и нарушению генеративной функции яичников. Более того, данные зарубежной литературы (C. Tamanini et al., 2003; V. D. Dixit, N. Parvizi, 2001; A. Honaramooz et al., 1999; K. M. Dhandopani, D. W. Brann, 2000) утверждают, что оксид азота включен в контроль секреции гонадотропин-рилизинг-гормона гипоталамусом, лютеинизирующего гормона гипофизом и контроль проявления половой охоты.
|
|
|
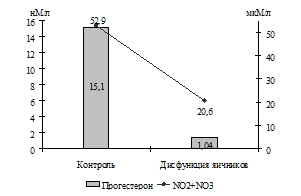
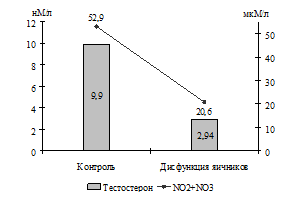
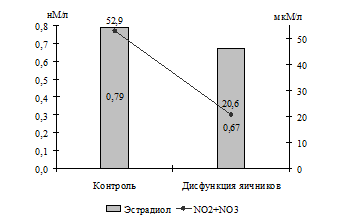
Рис. 1. Взаимосвязь содержания в крови коров половых стероидов и стабильных метаболитов оксида азота
Таким образом, в основе нарушения воспроизводительной функции и развития бесплодия у высокопродуктивных коров лежит расстройство в их организме эндокринных (гормональных) взаимоотношений, обмена белков, микроэлементов (селен, йод, марганец, медь), витаминов А и Е, сочетаемых с лактационной депрессией гипоталамо-гипофизарно-гонадальной системы и состоянием свободнорадикального окисления. При этом можно предположить, что такой свободный радикал, как оксид азота, является одним из компонентов многоуровневой системы регуляции репродуктивной функции млекопитающих (рис. 2).
Следовательно, одним из условий профилактики бесплодия и сохранения плодовитости высокопродуктивных коров должно стать устранение и предупреждение дисфункции эндокринной системы, адаптационных напряжений в обмене веществ и стрессовой дезадаптации организма животных за счет восполнения дефицита микроэлементов (в первую очередь селена и йода) и витаминов А и Е, выполняющих роль биокатализаторов всех метаболических процессов (В. Т. Самохин, 2003), ответственных за процессы пероксидации липидов и нормализацию синтеза простагландинов и стероидов (В. Т. Михайленко, Я., 1980; A. Agazwal et al., 2005; A. Sathya et al., 2007), а также включения в технологию ведения молочного скотоводства такого составляющего элемента, как применение средств стресс-корректорного действия.
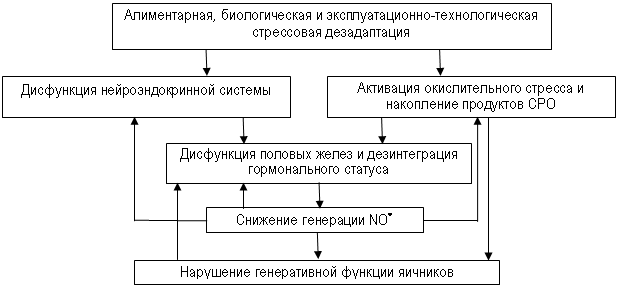
Рис.2. Гипотетическая схема развития дисфункции яичников у коров
3.5. Пероксидно-антиоксидантный статус коров при разном содержании в крови селена
Исследования проведены на животных, содержащихся в агробиоценозах с достаточным количеством селена (первая группа) и в зоне биогеохимического дефицита селена (вторая группа). Установлено, что пониженное содержание селена в биогеоценозе обусловливает и его дефицит в организме животных. Как следует из данных в таблице 10, содержание селена в крови коров второй группы составило от уровня первой группы в 2 месяца беременности 62,5% (Р<0,05), в 5 месяцев – 44,2% (Р<0,001), в 8,5 месяцев – 53,5% (Р<0,001).
Таблица 10
Показатели системы ПОЛ-АОЗ у беременных коров с разной
обеспеченностью селеном
Показатель | Беременность, мес | ||
2 | 5 | 8,5 | |
Селен, мкг% | 16,0±0,87 10,0±2,44 | 17,2±1,16 7,6±0,82 | 14,4±0,79 7,7±1,14 |
МДА, мкМ/л | 1,34±0,09 2,62±0,12 | 1,58±0,08 1,90±0,11 | 1,83±0,05 2,55±0,26 |
ГПО, мМ G-SH/л´мин | 11,7±0,75 8,2±0,82 | 10,3±0,30 10,0±0,45 | 15,8±0,93 11,1±0,52 |
ГР, мкМ G-SS-G/ л´мин | 273,0±8,51 143,0±6,51 | 267,0±15,22 144,6±4,40 | 360,0±8,44 144,0±6,81 |
СОД, усл. ед./мг Hb | 0,92±0,04 0,92±0,06 | 0,81±0,04 0,86±0,06 | 1,18±0,05 1,06±0,05 |
Каталаза, мМ Н2О2/л´мин | 34,9±1,16 33,2±1,68 | 36,0±0,96 32,4±1,04 | 38,7±0,74 37,0±1,01 |
Примечание: числитель – показатели коров I группы, знаменатель – показатели коров II группы.
Активизация процессов ПОЛ в динамике беременности у коров первой группы сопровождалась повышением в крови концентрации МДА к пяти месяцам стельности на 17,9% и к 8,5 – на 36,6% (Р<0,001). Одновременно с этим отмечено статистически достоверное (Р<0,01-0,001) компенсаторное увеличение активности глутатионпероксидазы на 35,4%, глуатионредуктазы – на 31,9%, супероксиддисмутазы – на 28,3%, каталазы – на 10,9%, что отражает активизацию ферментативного звена АОЗ, направленную на поддержание кислотно-основного гомеостаза организма. Максимальное повышение в этот период активности ГПО и ГР с одновременным значительным снижением содержания селена в крови свидетельствует об активном включении в систему антиоксидантной защиты глутатионового звена.
У коров второй группы отмечается более высокая интенсивность ПОЛ. Концентрация МДА в крови в два месяца беременности была выше на 95,5% (Р<0,001), в пять месяцев – на 20,2% (Р<0,05) и в восемь – на 39% (Р<0,05), чем у коров первой группы. В то же время активность ГПО у них в период глубокой стельности была ниже на 42,3% (Р<0,01), ГР – в 2,5 раза (Р<0,001), что свидетельствует о дисбалансе в состоянии ферментативного звена антиоксидантной системы у животных с дефицитом селена.
Со снятием фазы предродового и родового стрессового напряжения концентрация в крови коров первой группы МДА при нормальном течении послеродового периода уменьшилась на 83% (Р<0,001), а второй – на 32,8% (Р<0,05) – таблица 11, что отражает резкое падение интенсивности ПОЛ. Соответственно этому изменилось и состояние системы АОЗ. В крови коров первой группы активность ГПО снизилась на 67,9%, ГР – на 22,8%, СОД – на 63,9%, каталазы – на 49,4% (Р<0,01), а количество селена увеличилось на 11,9%, достигнув первоначальных значений. В крови коров второй группы изменения в активности ГПО составили 7,8%, ГР – 15,2%, СОД – 37,7%, каталазы – 10,1% и содержании селена – 70,1%.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 |
 Основные порталы (построено редакторами)
Основные порталы (построено редакторами)