

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
99-102. Crataegus ceratocarpa Kossych,
C. pojarkovae Kossych, C. sphaenophylla Pojark., C. stankovii Kossych
103. Potentilla depressa Willd. ex Schlecht.
104. P. taurica Willd. ex Schlecht.
105. Sorbus tauricola Zaikonn.
Rubiaceae
106. Asperula supina M. bsp. caespitans (Juz.) Pjatunina *
107. Galium xeroticum (Klokov) Soó
Santalaceae
108. Thesium krymense Romo, Didukh et Boratyński *
Saxifragaceae
109. Saxifraga irrigua M. Bieb.
Scrophulariaceae
110. Euphrasia taurica Ganesch. ex Popl.
111. Scrophularia exilis Popl.
112. S. goldeana Juz.
113-115. Veronica incana bsp. hololeuca (Juz.) A. Jelen.*; V. taurica bsp. bordzilowskii (Juz.) A. Jelen.*, subsp. taurica*
Thymelaeaceae
116. Daphne taurica Kotov
Valerianaceae
117. Valerianella falconida Schvedtsch.
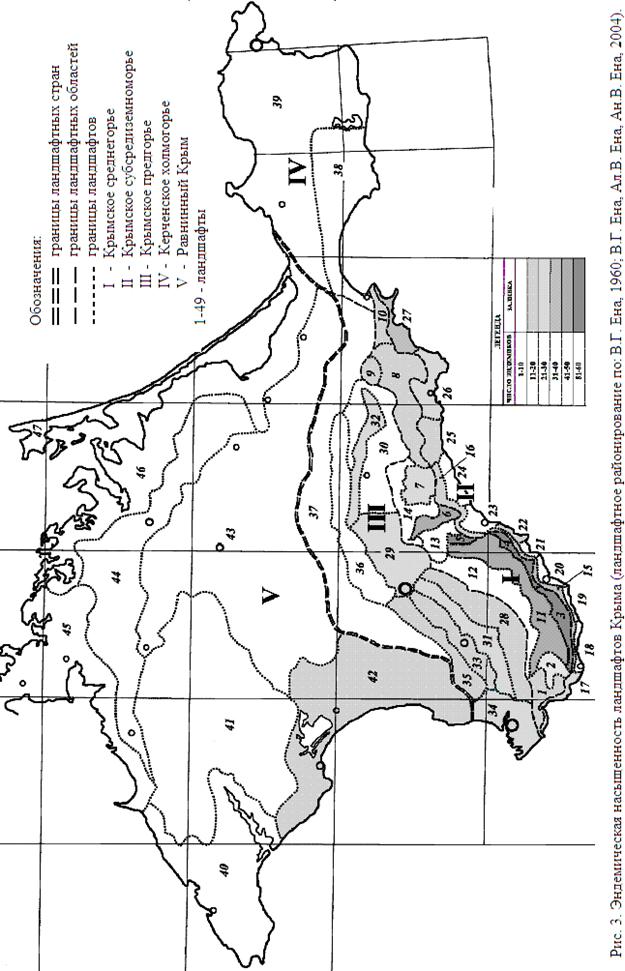
4.3. Фитониши и экониши крымского эндемизма. Большинство исследователей флоры Крыма (Стевен, 1856; Привалова, 1958; Рубцов, 1959; Шеляг-Сосонко и Дидух, 1980; Дидух, 1992; Рыфф, 1998, 2004 и др.) отмечали приуроченность региональных эндемиков к горной части полуострова, и прежде всего к яйлам. Проведенный нами анализ распределения крымского эндемизма в ландшафтах полуострова (согласно ландшафтному районированию: Ена, 1960; Ена, Ена, Ена, 2004) продемонстрировал, что они вмещают от 1 до 57 эндемичных видов (рис. 3). Наибольший уровень эндемизма, превышающий 50 видов, имеют западнояйлинские карстовые ландшафты области Главной Крымской гряды (номера ландшафтов даны в скобках) – Байдаро-Айпетринский (№3), Бабуган-Ялтинский (№4) и Чатырдагский (№5). В ландшафтах восточных яйл число эндемиков снижается: Демерджи-Долгоруковский (№6) – 41 вид, Караби-яйлинский (№7) – 27 видов и Агармышский останцево-яйлинский (9) – 17 видов; это напоминает ситуацию со снижением уровня эндемизма в том же географическом направлении среди внешнекуэстовых ландшафтов Крымского Предгорья. Подобным же образом снижается и число уникальных для каждого ландшафта локальных эндемиков – с 3-х в Байдаро-Айпетринском (№3) до 2-х в Бабуган-Ялтинском (№4) и по одному в Чатырдагском (№5), Демерджи-Долгоруковском (№6) и Караби-яйлинском (№7) ландшафтах. Высокий эндемизм на яйлах обеспечен их древним геологическим возрастом, изолированным положением, преимущественной безлесностью, огромным разнообразием геоморфологических, экотопических и ценотических проявлений. Подъяйлинские ландшафты обоих склонов Главной гряды также богаты эндемиками. Особенно их много в Северо-западном склоновом (№11) и Юго-западном крутосклонном (№15) подъяйлинских ландшафтах (соответственно 50 и 34 видов). Юго-восточный (№16) и Северо-восточный (№14) склоновые подъяйлинские ландшафты оказываются уже значительно обеднены эндемиками (соответственно 15 и 6 видов). Со снижением абсолютной высоты ландшафтов Главной гряды снижается и уровень эндемизма. В геологически самых молодых и геоморфологически однородных, наименее расчлененных ландшафтах с небольшим набором экотопов и низким фиторазнообразием в Равнинно-степном Крыму и Керченском степном холмогорье присутствуют единичные эндемики. Эндемическая насыщенность ландшафтов в целом возрастает в Крыму с севера на юг и с востока на запад (однако в Крымском Субсредиземноморье – с запада на восток) и с очевидностью положительно связана с увеличением геологического возраста, абсолютной высоты и сложности структуры ландшафтов.
Ландшафтно-экологическая специфика узких крымских эндемиков детально раскрыта нами на примере исследования двух модельных видов – Silene jailensis и Lepidium turczaninowii.
Что касается флористического районирования Крыма, то существующие схемы (Тахтаджян, 1978; Ан. А. Федоров, 1979; Заверуха, 1985) не могут в свете наших новых данных признаваться удовлетворительными. Отталкиваясь от этих схем, а также от принципов «Протокола флористического районирования», на основе изучения ареалов региональных эндемиков мы впервые осуществили картографическое флористическое районирование Крымского полуострова до районов включительно (рис. 4).
Эндемизм флоры Равнинного Крыма и Керченского полуострова ранее (Дидух, Вакаренко, 1987; Молчанов и др., 1988; Новосад, 1992, 1999; Голубев, 1996, 1999; Белянина, Шатко, 1999) сильно преувеличивался. По нашим данным, в первом случае есть только три локальных эндемика (Alyssum kotovii, Artemisia dzevanovskyi, Taraxacum salsum), а во втором – ни одного. Единичные местонахождения других крымских эндемиков и трансгрессоры из их числа (Anthemis dubia, Dianthus marschallii, Thymus dzevanovskyi и др.) при районировании в расчет не берутся. По той же причине такие точечные центры фиторазнообразия, как Джангуль, Казантип и Опук, не играют никакой хориономической роли.
Мы рассматриваем Крымско-Новороссийскую фитохорию как подпровинцию, а ее западную часть разделяем на три округа. Присвоение таких рангов объясняется, во-первых, общей комплексностью горнокрымской флоры, во-вторых, наличием нескольких хорологических комбинаций эндемичных видов; в–третьих, необходимостью приведения системы фитохорий в соответствие с «Протоколом флористического районирования».
Таким образом, в нашем видении Крым занимает следующее место в системе фитохорий Голарктического царства:
БОРЕАЛЬНОЕ ПОДЦАРСТВО
Циркумбореальная область
Восточноевропейская провинция
Понтическая подпровинция
Западнопонтический округ
Таврический р-н
Равниннокрымский подр-н
Керченский подр-н
ДРЕВНЕСРЕДИЗЕМНОМОРСКОЕ
ПОДЦАРСТВО
Средиземноморская область
Восточносредиземноморская подобласть
Эвксинская провинция
Крымско-Новороссийская подпровинция
Предгорнокрымский округ
Предгорнокрымский р-н
Восточногорнокрымский р-н
Яйлинский округ
Западнояйлинский р-н
Восточнояйлинский р-н
Южнокрымский округ
Байдарский р-н
Крымско-Южнобережный р-н
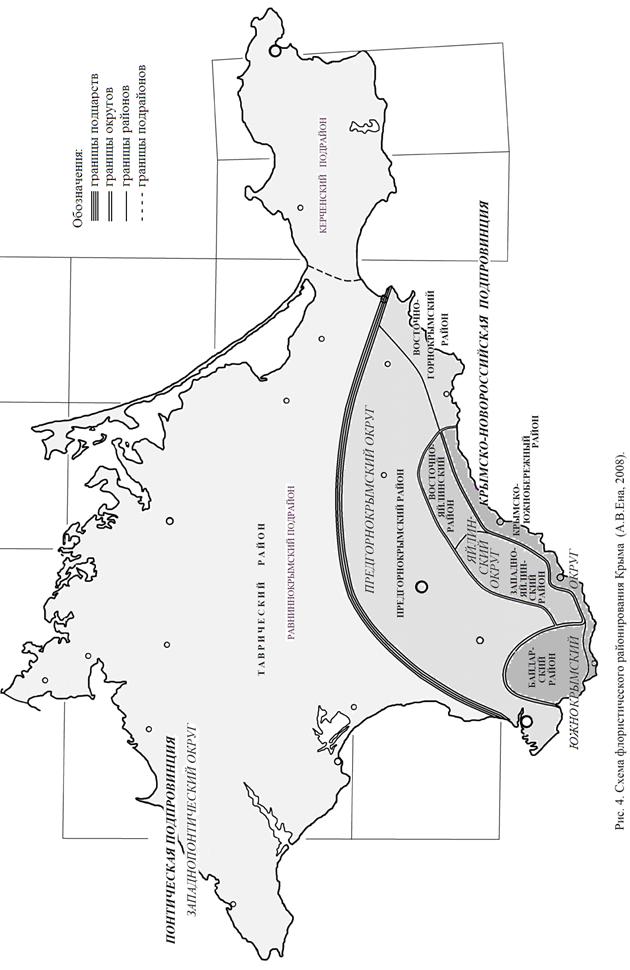
В нашей схеме восстанавливается средиземноморское понимание Эвксинской провинции, восходящее к В. П. Малееву (1940). Мы, в частности, приняли во внимание уточненные ареалы ряда видов, важная хориономическая роль которых прояснилась лишь в последнее время: Arbutus andrachne L., Cercis siliquastrum L., Cyclamen coum Mill., Heracleum pubescens (Hoffm.) M. Bieb., Himantoglossum caprinum (M. Bieb.) K. Koch,, Pinus brutia Ten., Ruscus ponticus Woronow ex Grossh. (Велчев и др., 1989; Ена, 1990; Паллас, 1999; Yena et al., 2005; Yena, Kraynyuk, 2007), а также правило непрерывности фитохорий (Толмачев, 1974).
раздел 5. Общие закономерности флористического эндемизма
5.1. Эволюция и эндемизм. Обсуждаются альтернативные модели видообразования – градуальная и сальтационная, кладистическая (возможный пример из крымских эндемиков: Veronica taurica subsp. taurica и subsp. bordzilowskii) и анагенетическая (возможный пример: Rumia crithmifolia). Показано, что это равно распространенные способы возникновения узких эндемиков с различными последствиями для их фито - и экониш.
Еще в одной альтернативной паре микроэволюционных гипотез – монотопической и политопической, как показывают многочисленные исследования (Willis, 1922; Cain, 1971; Попов, 1983; Вавилов, 1987; Махнев, 1987; Ней, Кумар, 2004 и др.), жизнеспособна лишь первая, обозначенная еще Ч. Дарвином (1991) как закон единства места происхождения вида. Признание совокупности политопически возникших групп растений единым видом проблематично прежде всего из-за неповторимости генетических комбинаций. На основании дарвиновского закона о монотопическом происхождении видов мы считаем, что в принципе все виды растений на заре своего существования были узкими эндемиками. Из приведенных соображений вытекает, что узкий эндемизм является облигатной стадией в становлении каждого молодого вида, в т. ч. аллопатрического, симпатрического и гибридного происхождения. В свете сказанного совершенно по-новому предстает сама проблема эндемизма, который, по сути, получает не эпизодическую, а одну из ключевых ролей во всемирной драме эволюции растительного покрова. Отсюда проясняется весомая причина, по которой узкие эндемики заслуживают абсолютного приоритета при изучении и сохранении фиторазнообразия: они не просто редкие или оригинальные формы, а страховой потенциал преемственности и непрерывности эволюционных изменений во флоре. Уничтожение эндемиков подрывает стабильность флорогенеза и освобождает путь инвайдерам. Подобно превращению редкого на родине вида в агрессивного инвайдера на чужбине, существует событийный тренд, позволяющий при определенных условиях узкому эндемику резко расширить ареал (Haselwood, Motter, 1983; Strid, 1989; Silva, 1999 и др.). С другой стороны, выработаны подходы, позволяющие предсказывать появление новых эндемиков (Gottlieb, 1979; Loveless, Hamrick, 1988; Fréville et al., 1998 и др.), и они могут быть экстраполированы по крайней мере на некоторые из крымских эндемичных таксонов, например, из р. Centaurea, Cirsium и Anthemis.
5.2. Эндемики среди конгенеров. Далеко не все редкие эндемики имеют узкий диапазон толерантности, так что узкие эндемики и более обычные конгенеры часто одинаково успешно развиваются ex situ. Для выяснения причин несоответствия ясно ограниченной фитониши и потенциально широкой экониши узких эндемиков в последние годы проведены специальные исследования пар конгенеров – эндемика и андемика из одного рода – по десяткам различных признаков, in situ et ex situ (Baskin et al., 1997; Radford et al., 2000; Lavergne et al., 2004; Kephart, 2005 и др.). Во всех случаях в основе склонности эндемиков к сохранению размеров ареала лежит не только их имманентная неконкурентоспособность, но также влияние консортов на генеративную сферу растений, что в сочетании с особенностями цветка приводит к низкой семенной продуктивности in situ. Подобных отечественных исследований пока нет, но мы считаем, что данный вывод подтверждается крымским примером с локальным эндемиком Crataegus pojarkovae, который, в отличие от обычного конгенера C. orientalis Pall. ex M. Bieb., имеет крайне низкую всхожесть семян – 3-4% (Косых, 1967) и высокую стерильность пыльцы – 23,9% (Летухова, 2003), а самое главное, паразитический гриб Gymnosporangium clavarieforme (Jacq.) DC., встречающийся у C. orientalis только на листьях, у C. pojarkovae поражает плоды (Исиков, Шевченко, 1991).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 |
